

A Kinase-Independent Role for Cyclin-Dependent Kinase 19 in p53 Response
- PMID: 28416637
- PMCID: PMC5472832
- DOI: 10.1128/MCB.00626-16
A Kinase-Independent Role for Cyclin-Dependent Kinase 19 in p53 Response
Abstract
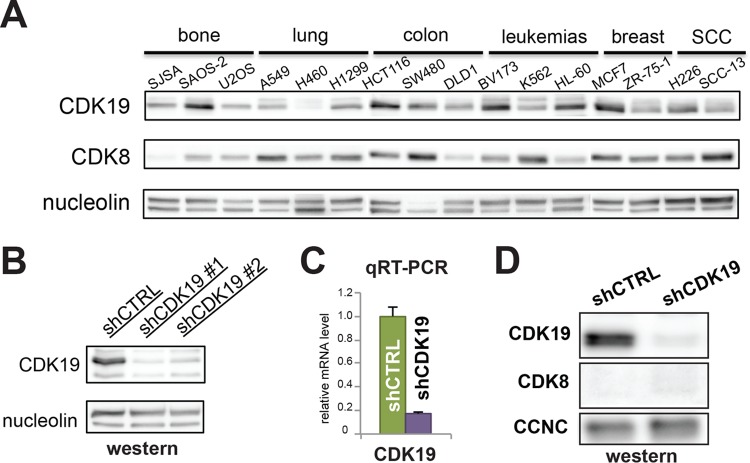
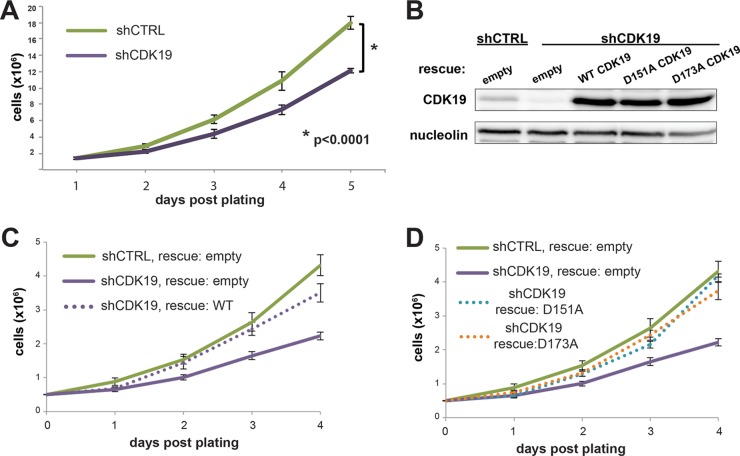
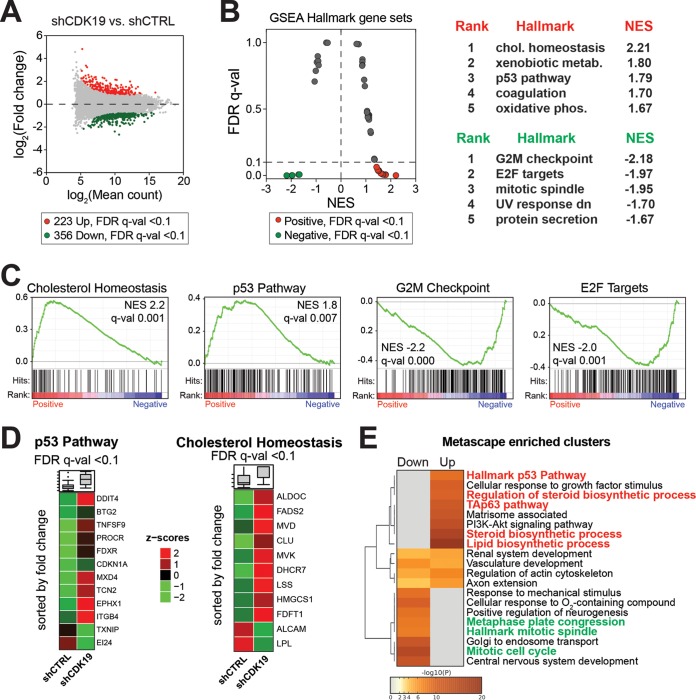
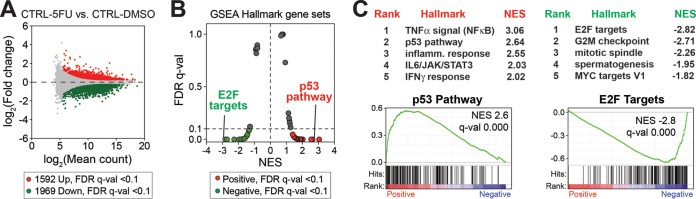
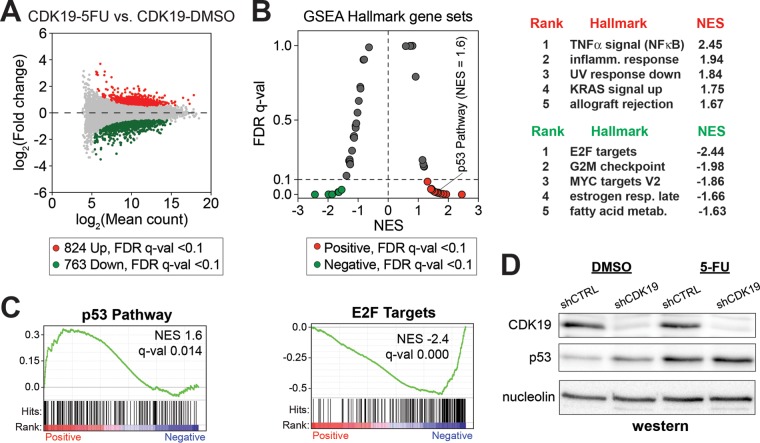
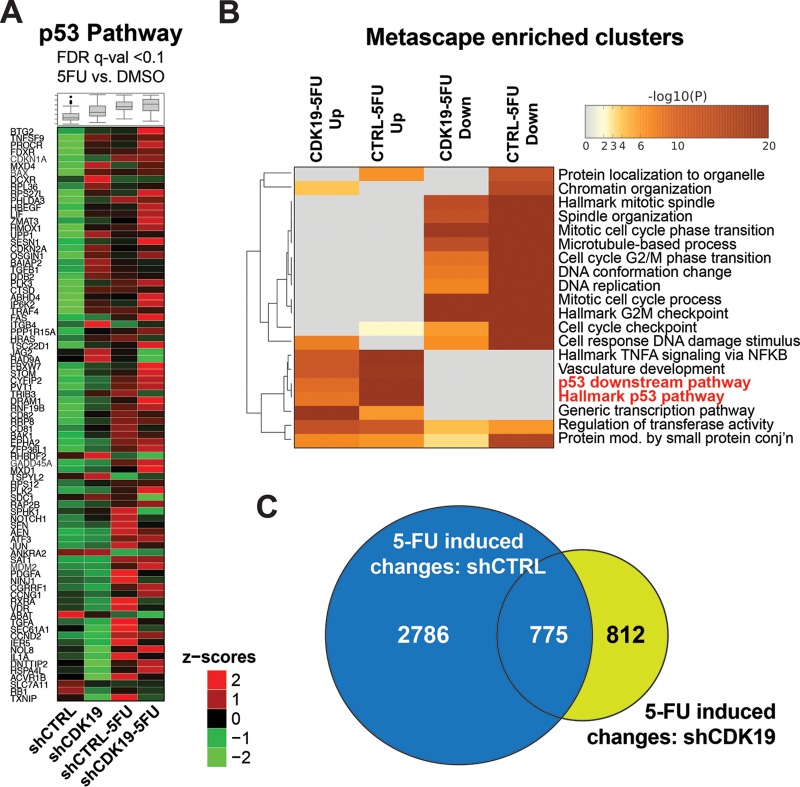
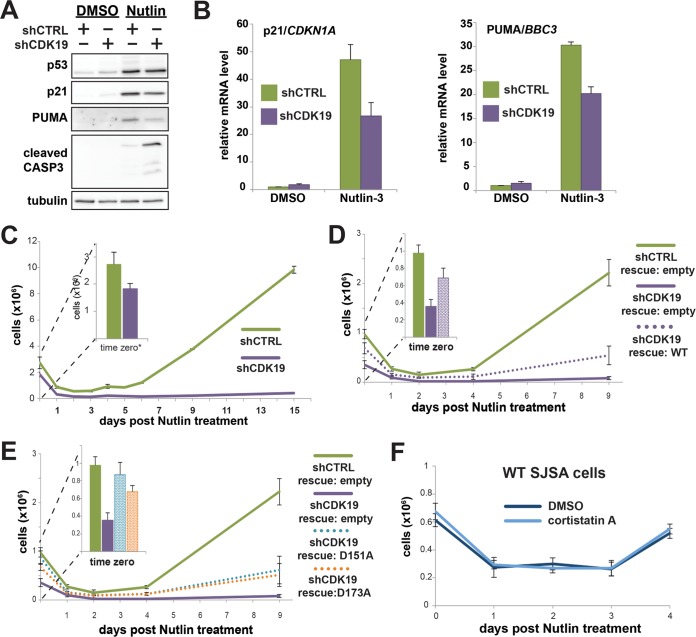
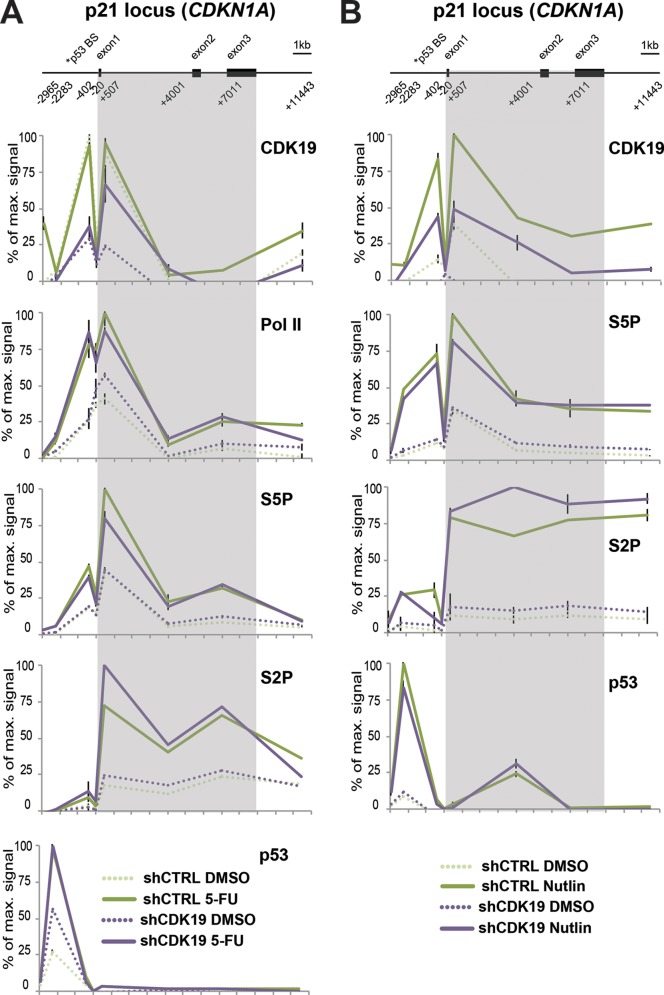
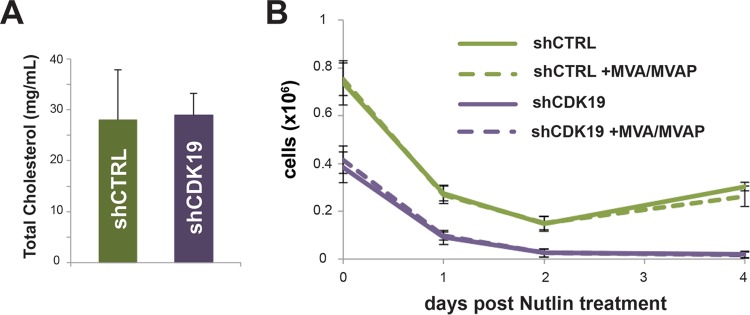
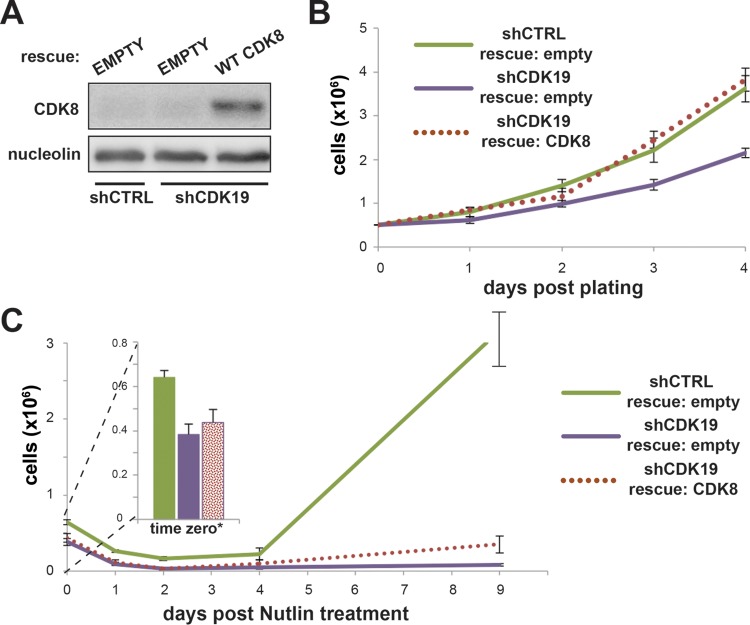
The human Mediator complex regulates RNA polymerase II transcription genome-wide. A general factor that regulates Mediator function is the four-subunit kinase module, which contains either cyclin-dependent kinase 8 (CDK8) or CDK19. Whereas CDK8 is linked to specific signaling cascades and oncogenesis, the cellular roles of its paralog, CDK19, are poorly studied. We discovered that osteosarcoma cells (SJSA) are naturally depleted of CDK8 protein. Whereas stable CDK19 knockdown was tolerated in SJSA cells, proliferation was reduced. Notably, proliferation defects were rescued upon the reexpression of wild-type or kinase-dead CDK19. Comparative RNA sequencing analyses showed reduced expression of mitotic genes and activation of genes associated with cholesterol metabolism and the p53 pathway in CDK19 knockdown cells. SJSA cells treated with 5-fluorouracil, which induces metabolic and genotoxic stress and activates p53, further implicated CDK19 in p53 target gene expression. To better probe the p53 response, SJSA cells (shCDK19 versus shCTRL) were treated with the p53 activator nutlin-3. Remarkably, CDK19 was required for SJSA cells to return to a proliferative state after nutlin-3 treatment, and this effect was kinase independent. These results implicate CDK19 as a regulator of p53 stress responses and suggest a role for CDK19 in cellular resistance to nutlin-3.
Keywords: 5-FU; 5-fluorouracil; CDK19; CDK8; Mediator kinase; RNA-Seq; SJSA; cholesterol; cortistatin A; drug resistance; nutlin; nutlin-3; osteosarcoma; p53; stress; transcription.
Copyright © 2017 Audetat et al.
Figures
Similar articles
-
Oncogenic exon 2 mutations in Mediator subunit MED12 disrupt allosteric activation of cyclin C-CDK8/19.J Biol Chem. 2018 Mar 30;293(13):4870-4882. doi: 10.1074/jbc.RA118.001725. Epub 2018 Feb 13. J Biol Chem. 2018. PMID: 29440396 Free PMC article.
-
Identification of target genes for the CDK subunits of the Mediator complex.Genes Cells. 2011 Dec;16(12):1208-18. doi: 10.1111/j.1365-2443.2011.01565.x. Genes Cells. 2011. PMID: 22117896
-
HDM2 antagonist Nutlin-3 disrupts p73-HDM2 binding and enhances p73 function.Oncogene. 2008 Feb 7;27(7):997-1003. doi: 10.1038/sj.onc.1210707. Epub 2007 Aug 13. Oncogene. 2008. PMID: 17700533
-
CDK8: a positive regulator of transcription.Transcription. 2010 Jul-Aug;1(1):4-12. doi: 10.4161/trns.1.1.12373. Transcription. 2010. PMID: 21327159 Free PMC article. Review.
-
The Mediator kinase module: an interface between cell signaling and transcription.Trends Biochem Sci. 2022 Apr;47(4):314-327. doi: 10.1016/j.tibs.2022.01.002. Epub 2022 Feb 19. Trends Biochem Sci. 2022. PMID: 35193797 Free PMC article. Review.
Cited by
-
Gene signatures of cyclin-dependent kinases: a comparative study in naïve early and advanced stages of lung metastasis breast cancer among pre- and post-menopausal women.Genes Cancer. 2021 Feb 10;12:1-11. doi: 10.18632/genesandcancer.209. eCollection 2021. Genes Cancer. 2021. PMID: 33868579 Free PMC article.
-
Understanding Obesity as a Risk Factor for Uterine Tumors Using Drosophila.Adv Exp Med Biol. 2019;1167:129-155. doi: 10.1007/978-3-030-23629-8_8. Adv Exp Med Biol. 2019. PMID: 31520353 Review.
-
Mediator kinase inhibition reverses castration resistance of advanced prostate cancer.J Clin Invest. 2024 Mar 28;134(10):e176709. doi: 10.1172/JCI176709. J Clin Invest. 2024. PMID: 38546787 Free PMC article.
-
Rethink of EGFR in Cancer With Its Kinase Independent Function on Board.Front Oncol. 2019 Aug 23;9:800. doi: 10.3389/fonc.2019.00800. eCollection 2019. Front Oncol. 2019. PMID: 31508364 Free PMC article.
-
Suppression of p53 response by targeting p53-Mediator binding with a stapled peptide.Cell Rep. 2022 Apr 5;39(1):110630. doi: 10.1016/j.celrep.2022.110630. Cell Rep. 2022. PMID: 35385747 Free PMC article.
References
-
- Daniels DL, Ford M, Schwinn MK, Benink H, Galbraith MD, Amunugama R, Jones R, Allen D, Okazaki N, Yamakawa H, Miki F, Nagase T, Espinosa JM, Urh M. 2013. Mutual exclusivity of MED12/MED12L, MED13/13L, and CDK8/19 paralogs revealed within the CDK-Mediator kinase module. J Proteomics Bioinform S2:004.
-
- Pelish HE, Liau BB, Nitulescu II, Tangpeerachaikul A, Poss ZC, Da Silva DH, Caruso BT, Arefolov A, Fadeyi O, Christie AL, Du K, Banka D, Schneider EV, Jestel A, Zou G, Si C, Ebmeier CC, Bronson RT, Krivtsov AV, Myers AG, Kohl NE, Kung AL, Armstrong SA, Lemieux ME, Taatjes DJ, Shair MD. 2015. Mediator kinase inhibition further activates super-enhancer-associated genes in AML. Nature 526:273–276. doi:10.1038/nature14904. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous