

Domain-dependent effects of insulin and IGF-1 receptors on signalling and gene expression
- PMID: 28345670
- PMCID: PMC5378997
- DOI: 10.1038/ncomms14892
Domain-dependent effects of insulin and IGF-1 receptors on signalling and gene expression
Abstract
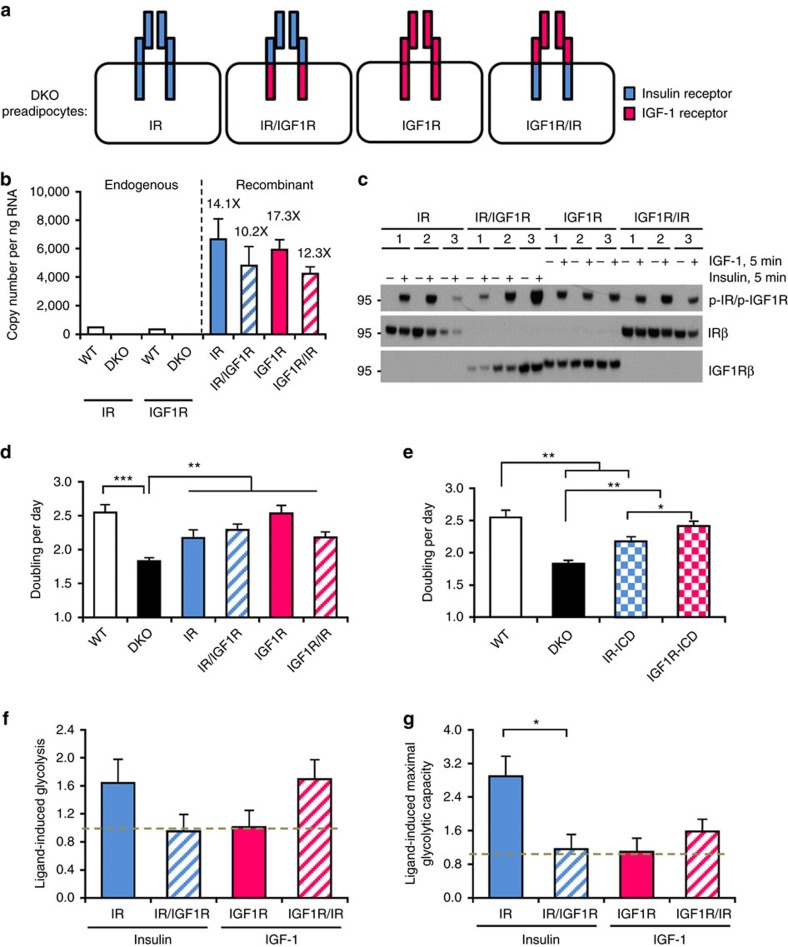
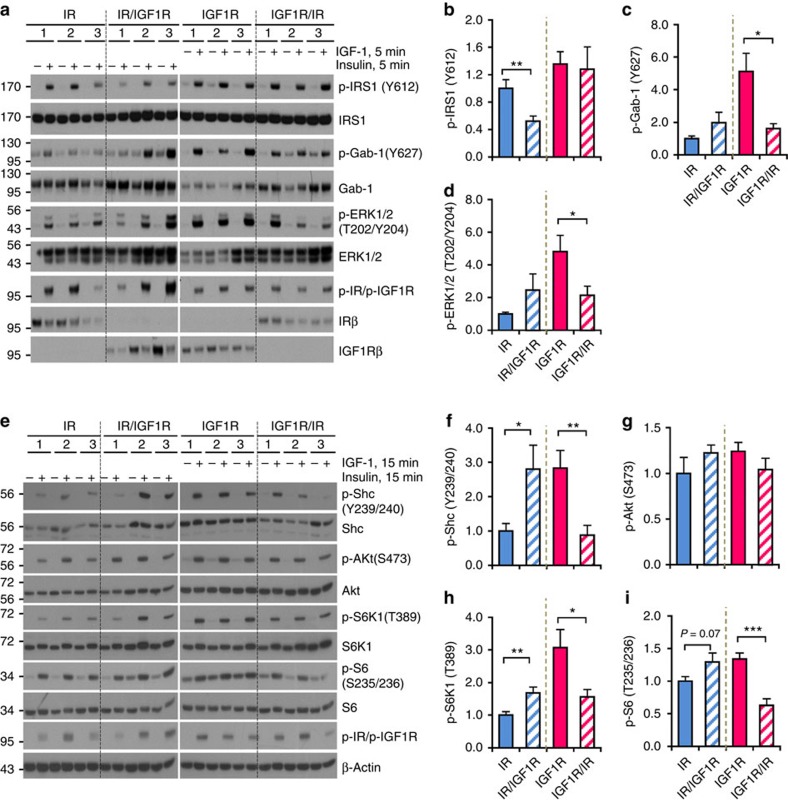
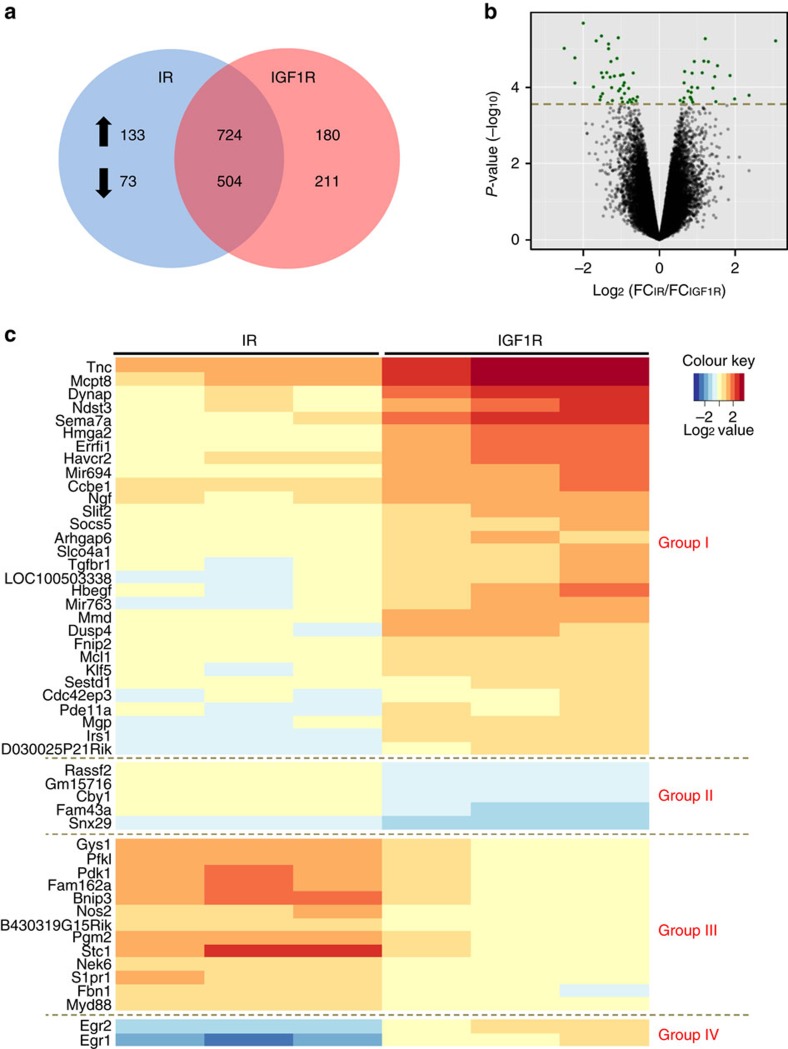
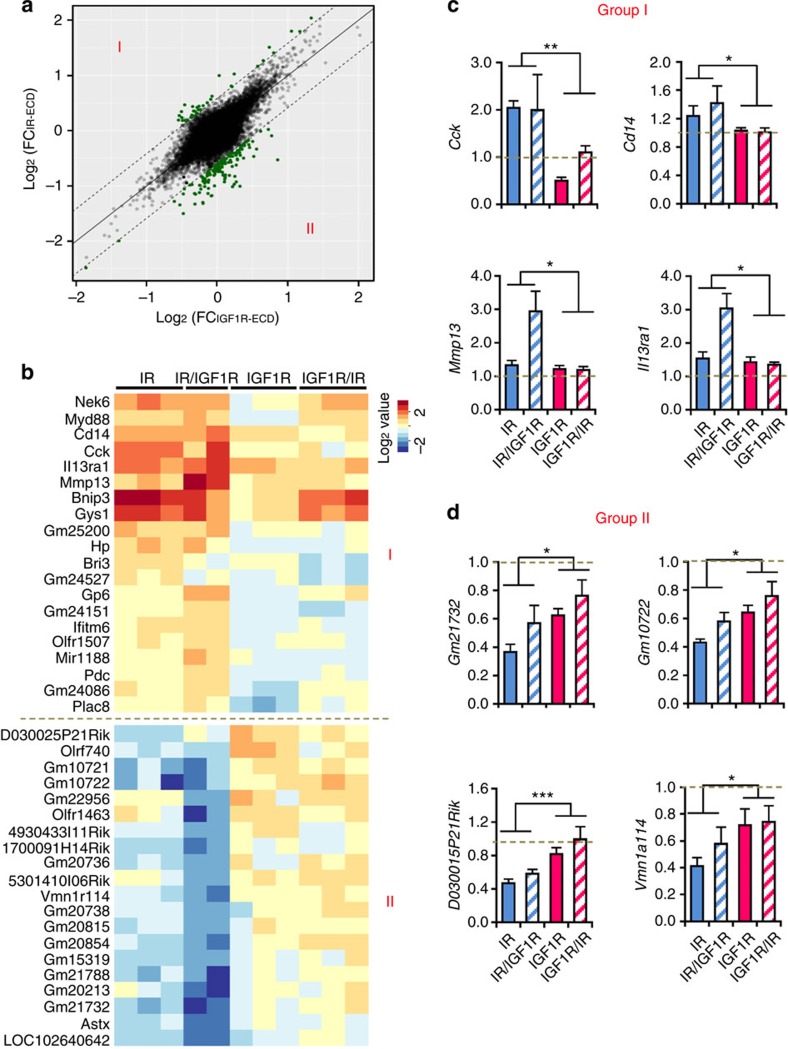
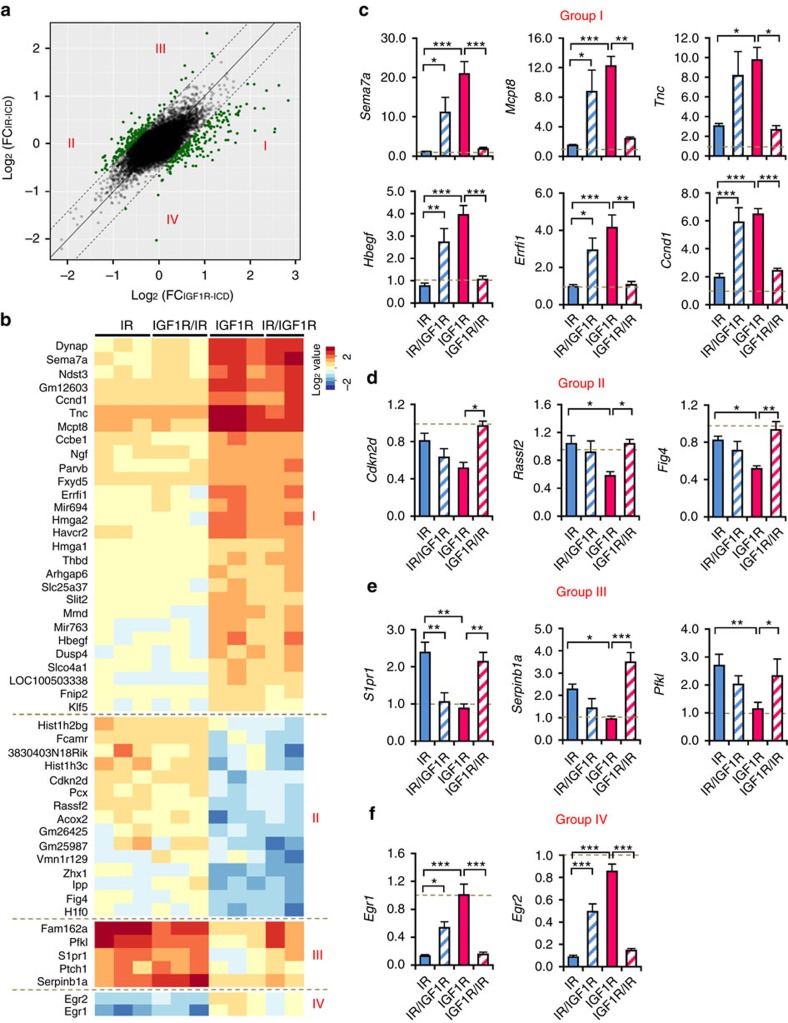
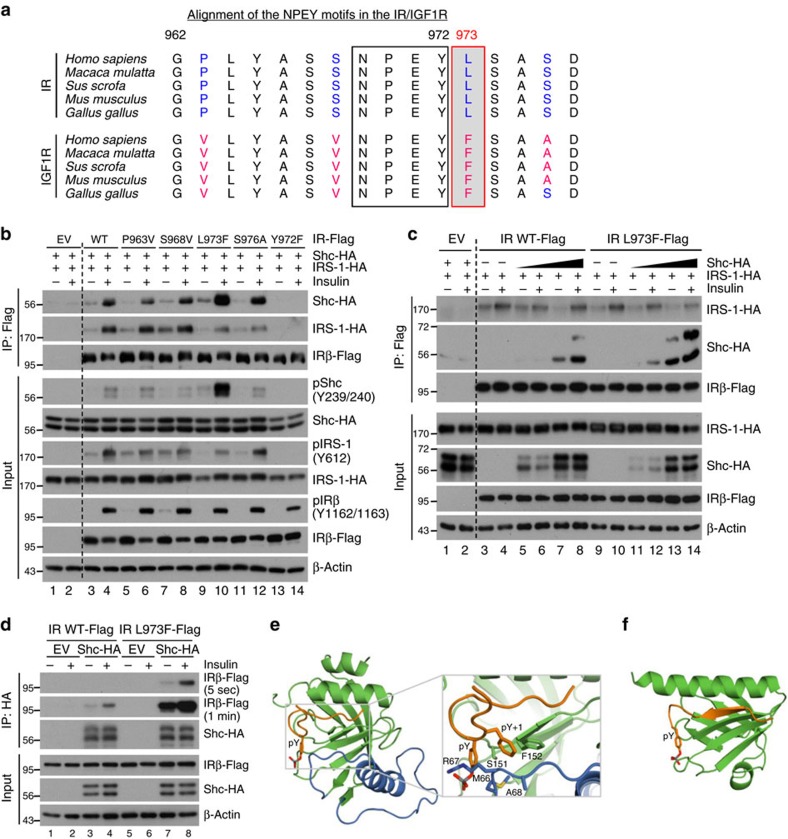
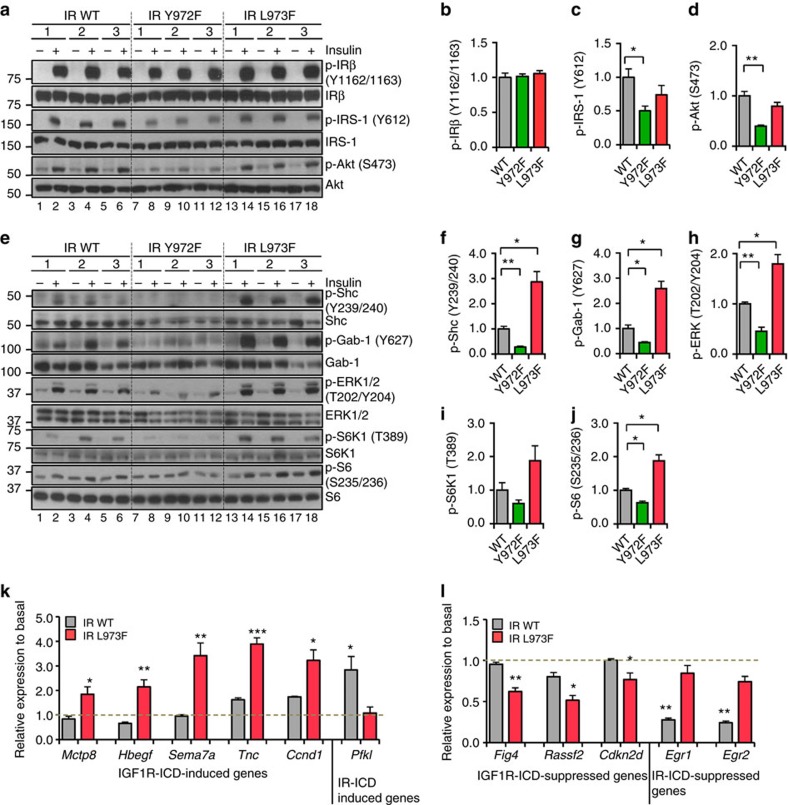
Despite a high degree of homology, insulin receptor (IR) and IGF-1 receptor (IGF1R) mediate distinct cellular and physiological functions. Here, we demonstrate how domain differences between IR and IGF1R contribute to the distinct functions of these receptors using chimeric and site-mutated receptors. Receptors with the intracellular domain of IGF1R show increased activation of Shc and Gab-1 and more potent regulation of genes involved in proliferation, corresponding to their higher mitogenic activity. Conversely, receptors with the intracellular domain of IR display higher IRS-1 phosphorylation, stronger regulation of genes in metabolic pathways and more dramatic glycolytic responses to hormonal stimulation. Strikingly, replacement of leucine973 in the juxtamembrane region of IR to phenylalanine, which is present in IGF1R, mimics many of these signalling and gene expression responses. Overall, we show that the distinct activities of the closely related IR and IGF1R are mediated by their intracellular juxtamembrane region and substrate binding to this region.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
Tyrosine residues in the C-terminal domain of the insulin-like growth factor-I receptor mediate mitogenic and tumorigenic signals.Endocrinology. 1997 Jul;138(7):2979-88. doi: 10.1210/endo.138.7.5281. Endocrinology. 1997. PMID: 9202243
-
Leucine-973 is a crucial residue differentiating insulin and IGF-1 receptor signaling.J Clin Invest. 2023 Feb 15;133(4):e161472. doi: 10.1172/JCI161472. J Clin Invest. 2023. PMID: 36548088 Free PMC article.
-
Tamoxifen interferes with the insulin-like growth factor I receptor (IGF-IR) signaling pathway in breast cancer cells.Cancer Res. 1997 Jul 1;57(13):2606-10. Cancer Res. 1997. PMID: 9205064
-
Differential regulation of signaling pathways for insulin and insulin-like growth factor I.Acta Biochim Pol. 1999;46(1):51-60. Acta Biochim Pol. 1999. PMID: 10453981 Review.
-
Role of estrogen receptor alpha in modulating IGF-I receptor signaling and function in breast cancer.J Exp Clin Cancer Res. 2004 Sep;23(3):385-94. J Exp Clin Cancer Res. 2004. PMID: 15595626 Review.
Cited by
-
Evaluation of the Effect of FOXO3 rs13217795 Genotype and Minor Allele (C) on Clinical Chemistry and Genetic Risk of Diabetes Among the Elderly Individuals from Northern India.Mol Syndromol. 2022 Feb;13(2):99-107. doi: 10.1159/000518636. Epub 2021 Oct 8. Mol Syndromol. 2022. PMID: 35418822 Free PMC article.
-
Therapeutic IGF-I receptor inhibition alters fibrocyte immune phenotype in thyroid-associated ophthalmopathy.Proc Natl Acad Sci U S A. 2021 Dec 28;118(52):e2114244118. doi: 10.1073/pnas.2114244118. Proc Natl Acad Sci U S A. 2021. PMID: 34949642 Free PMC article.
-
Rab5a activates IRS1 to coordinate IGF-AKT-mTOR signaling and myoblast differentiation during muscle regeneration.Cell Death Differ. 2020 Aug;27(8):2344-2362. doi: 10.1038/s41418-020-0508-1. Epub 2020 Feb 12. Cell Death Differ. 2020. PMID: 32051546 Free PMC article.
-
Multi-layered proteomics identifies insulin-induced upregulation of the EphA2 receptor via the ERK pathway which is dependent on low IGF1R level.Sci Rep. 2024 Nov 21;14(1):28856. doi: 10.1038/s41598-024-77817-5. Sci Rep. 2024. PMID: 39572596 Free PMC article.
-
Insulin-Like Growth Factor 1 Attenuates the Pro-Inflammatory Phenotype of Neutrophils in Myocardial Infarction.Front Immunol. 2022 Jul 15;13:908023. doi: 10.3389/fimmu.2022.908023. eCollection 2022. Front Immunol. 2022. PMID: 35911749 Free PMC article.
References
-
- Kahn C. R., Freychet P., Roth J. & Neville D. M. Jr Quantitative aspects of the insulin-receptor interaction in liver plasma membranes. J. Biol. Chem. 249, 2249–2257 (1974). - PubMed
-
- Steele-Perkins G. et al.. Expression and characterization of a functional human insulin-like growth factor I receptor. J. Biol. Chem. 263, 11486–11492 (1988). - PubMed
-
- Taniguchi C. M., Emanuelli B. & Kahn C. R. Critical nodes in signalling pathways: insights into insulin action. Nat. Rev. Mol. Cell Biol. 7, 85–96 (2006). - PubMed
-
- Accili D. et al.. Early neonatal death in mice homozygous for a null allele of the insulin receptor gene. Nat. Genet. 12, 106–109 (1996). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
