

Exposure to selenomethionine causes selenocysteine misincorporation and protein aggregation in Saccharomyces cerevisiae
- PMID: 28303947
- PMCID: PMC5355996
- DOI: 10.1038/srep44761
Exposure to selenomethionine causes selenocysteine misincorporation and protein aggregation in Saccharomyces cerevisiae
Abstract
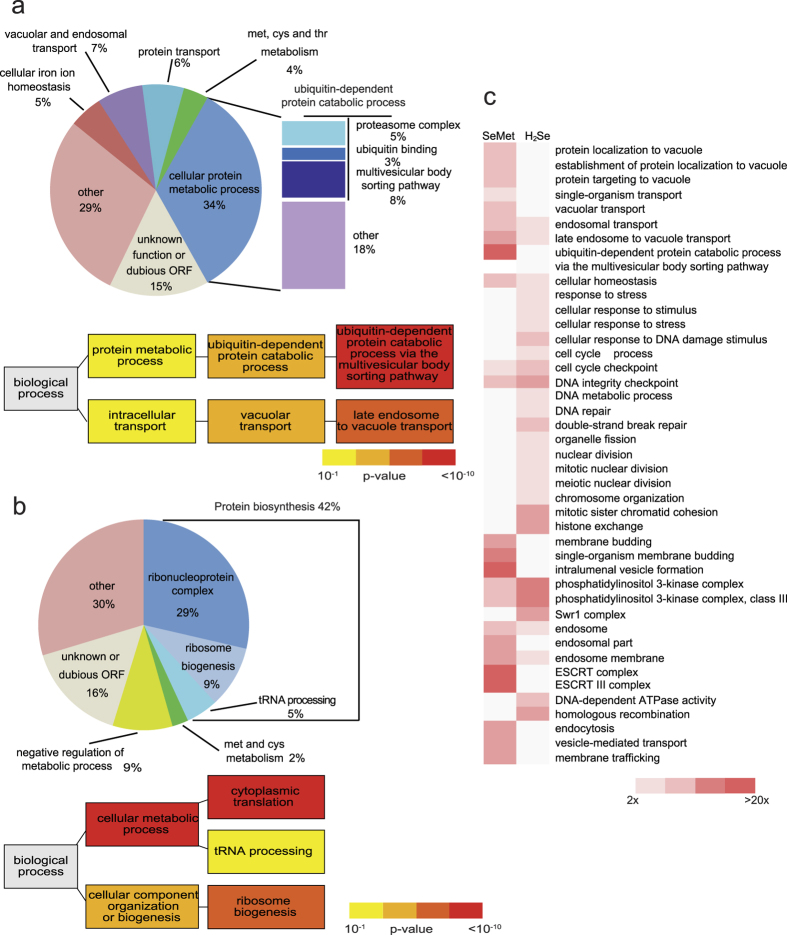
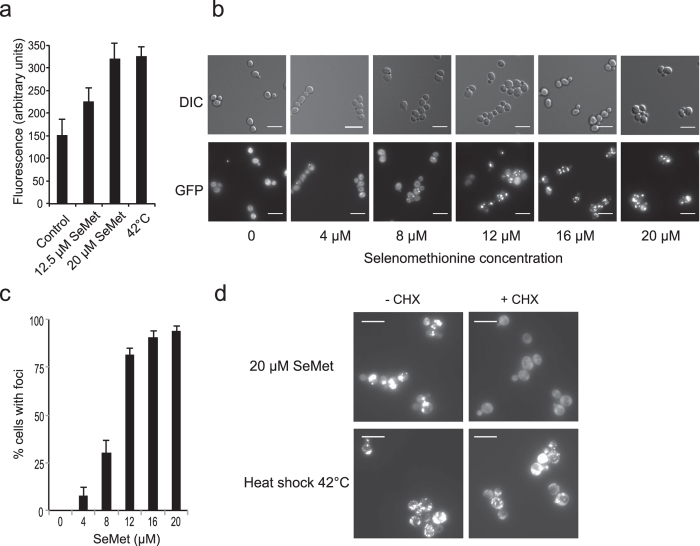
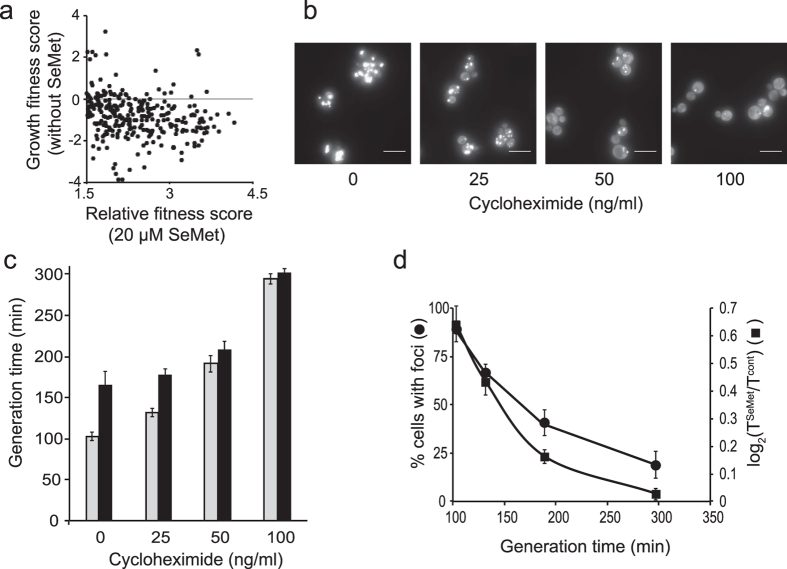
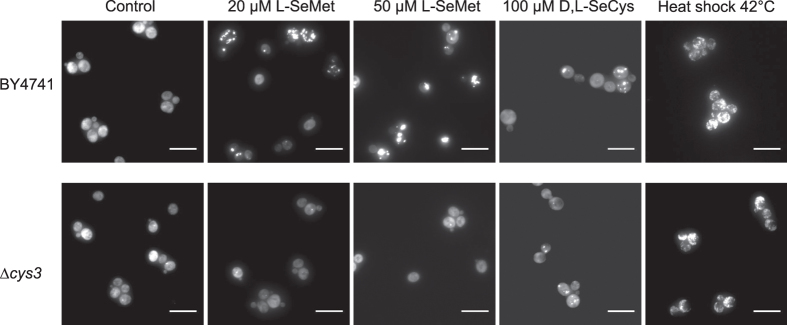
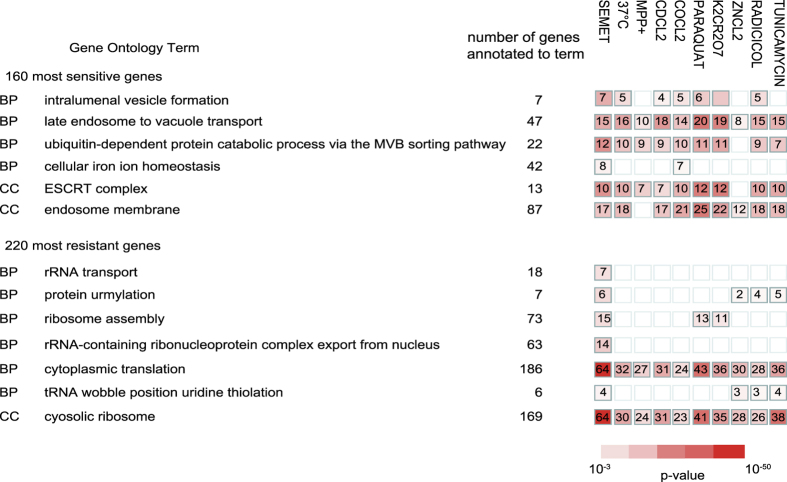
Selenomethionine, a dietary supplement with beneficial health effects, becomes toxic if taken in excess. To gain insight into the mechanisms of action of selenomethionine, we screened a collection of ≈5900 Saccharomyces cerevisiae mutants for sensitivity or resistance to growth-limiting amounts of the compound. Genes involved in protein degradation and synthesis were enriched in the obtained datasets, suggesting that selenomethionine causes a proteotoxic stress. We demonstrate that selenomethionine induces an accumulation of protein aggregates by a mechanism that requires de novo protein synthesis. Reduction of translation rates was accompanied by a decrease of protein aggregation and of selenomethionine toxicity. Protein aggregation was supressed in a ∆cys3 mutant unable to synthetize selenocysteine, suggesting that aggregation results from the metabolization of selenomethionine to selenocysteine followed by translational incorporation in the place of cysteine. In support of this mechanism, we were able to detect random substitutions of cysteinyl residues by selenocysteine in a reporter protein. Our results reveal a novel mechanism of toxicity that may have implications in higher eukaryotes.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
A cysteinyl-tRNA synthetase variant confers resistance against selenite toxicity and decreases selenocysteine misincorporation.J Biol Chem. 2019 Aug 23;294(34):12855-12865. doi: 10.1074/jbc.RA119.008219. Epub 2019 Jul 11. J Biol Chem. 2019. PMID: 31296657 Free PMC article.
-
Trans-sulfuration Pathway Seleno-amino Acids Are Mediators of Selenomethionine Toxicity in Saccharomyces cerevisiae.J Biol Chem. 2015 Apr 24;290(17):10741-50. doi: 10.1074/jbc.M115.640375. Epub 2015 Mar 5. J Biol Chem. 2015. PMID: 25745108 Free PMC article.
-
Recent advances in the mechanism of selenoamino acids toxicity in eukaryotic cells.Biomol Concepts. 2017 May 24;8(2):93-104. doi: 10.1515/bmc-2017-0007. Biomol Concepts. 2017. PMID: 28574376 Review.
-
Selenomethionine metabolism and its toxicity in yeast.Biomol Concepts. 2013 Dec;4(6):611-6. doi: 10.1515/bmc-2013-0033. Biomol Concepts. 2013. PMID: 25436761 Review.
-
Selective selC-independent selenocysteine incorporation into formate dehydrogenases.PLoS One. 2013 Apr 25;8(4):e61913. doi: 10.1371/journal.pone.0061913. Print 2013. PLoS One. 2013. PMID: 23634217 Free PMC article.
Cited by
-
The Role of Selenium in Arsenic and Cadmium Toxicity: an Updated Review of Scientific Literature.Biol Trace Elem Res. 2020 Jan;193(1):44-63. doi: 10.1007/s12011-019-01691-w. Epub 2019 Mar 15. Biol Trace Elem Res. 2020. PMID: 30877523 Free PMC article. Review.
-
Selenium in combination with a tomato lipid extract as a therapy for benign prostatic hyperplasia and its alterations in rats with induced BPH.J Cell Mol Med. 2023 Oct;27(20):3147-3156. doi: 10.1111/jcmm.17903. Epub 2023 Sep 19. J Cell Mol Med. 2023. PMID: 37726932 Free PMC article.
-
A cysteinyl-tRNA synthetase variant confers resistance against selenite toxicity and decreases selenocysteine misincorporation.J Biol Chem. 2019 Aug 23;294(34):12855-12865. doi: 10.1074/jbc.RA119.008219. Epub 2019 Jul 11. J Biol Chem. 2019. PMID: 31296657 Free PMC article.
-
SEXUAL DIMORPHISM IN SELENIUM METABOLISM AND SELENOPROTEINS.Free Radic Biol Med. 2018 Nov 1;127:198-205. doi: 10.1016/j.freeradbiomed.2018.03.036. Epub 2018 Mar 21. Free Radic Biol Med. 2018. PMID: 29572096 Free PMC article. Review.
-
Organic Selenium (OH-MetSe) Effect on Whole Body Fatty Acids and Mx Gene Expression against Viral Infection in Gilthead Seabream (Sparus aurata) Juveniles.Animals (Basel). 2021 Sep 30;11(10):2877. doi: 10.3390/ani11102877. Animals (Basel). 2021. PMID: 34679898 Free PMC article.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
