

Roles of MIWI, MILI and PLD6 in small RNA regulation in mouse growing oocytes
- PMID: 28115634
- PMCID: PMC5435931
- DOI: 10.1093/nar/gkx027
Roles of MIWI, MILI and PLD6 in small RNA regulation in mouse growing oocytes
Abstract
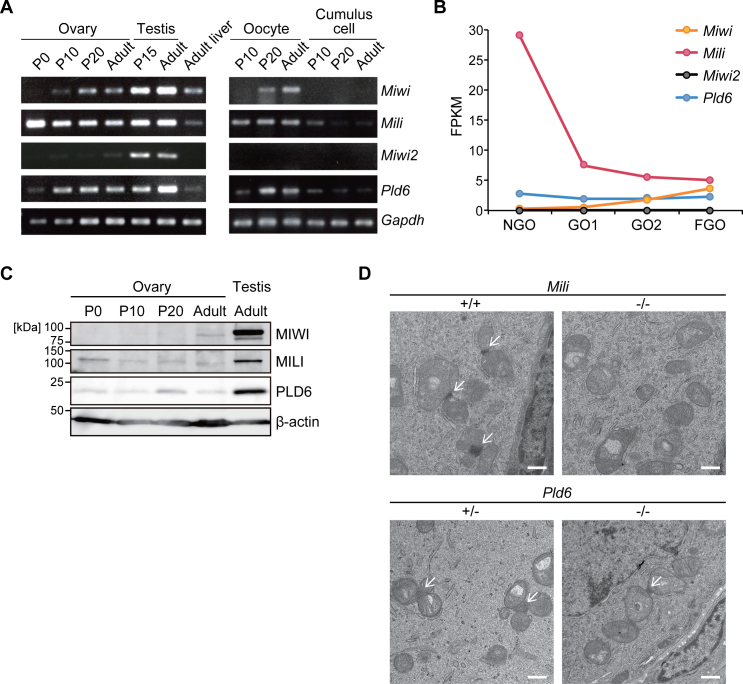
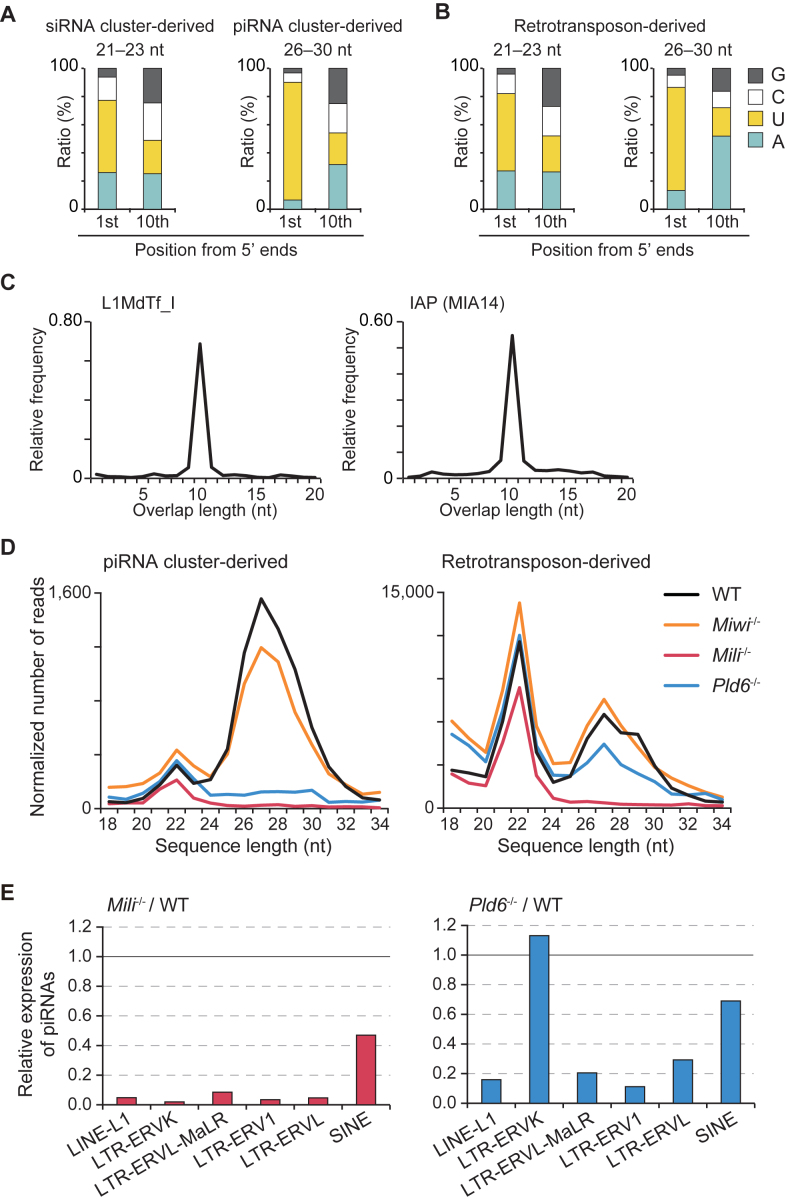
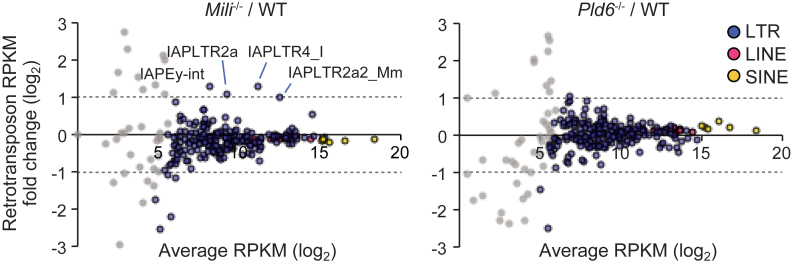
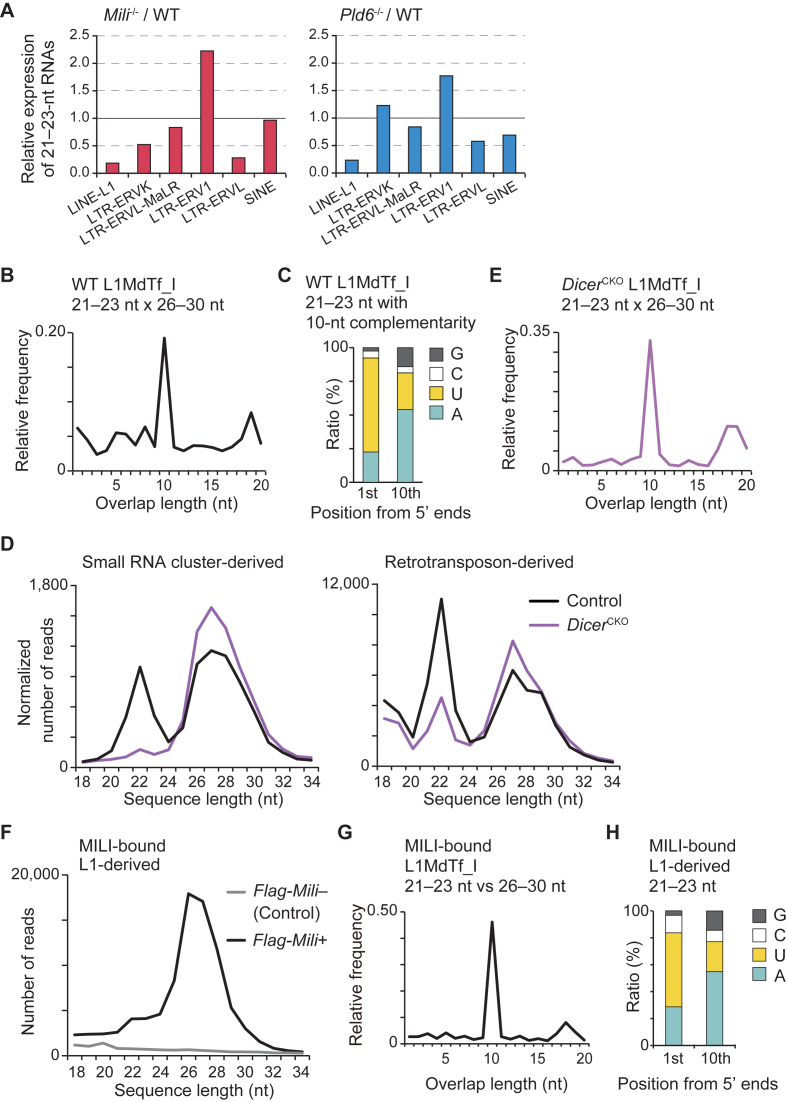
The mouse PIWI-interacting RNA (piRNA) pathway produces a class of 26-30-nucleotide (nt) small RNAs and is essential for spermatogenesis and retrotransposon repression. In oocytes, however, its regulation and function are poorly understood. In the present study, we investigated the consequences of loss of piRNA-pathway components in growing oocytes. When MILI (or PIWIL2), a PIWI family member, was depleted by gene knockout, almost all piRNAs disappeared. This severe loss of piRNA was accompanied by an increase in transcripts derived from specific retrotransposons, especially IAPs. MIWI (or PIWIL1) depletion had a smaller effect. In oocytes lacking PLD6 (or ZUCCHINI or MITOPLD), a mitochondrial nuclease/phospholipase involved in piRNA biogenesis in male germ cells, the piRNA level was decreased to 50% compared to wild-type, a phenotype much milder than that in males. Since PLD6 is essential for the creation of the 5΄ ends of primary piRNAs in males, the presence of mature piRNA in PLD6-depleted oocytes suggests the presence of compensating enzymes. Furthermore, we identified novel 21-23-nt small RNAs, termed spiRNAs, possessing a 10-nt complementarity with piRNAs, which were produced dependent on MILI and independent of DICER. Our study revealed the differences in the biogenesis and function of the piRNA pathway between sexes.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
Mitochondrial membrane-based initial separation of MIWI and MILI functions during pachytene piRNA biogenesis.Nucleic Acids Res. 2019 Mar 18;47(5):2594-2608. doi: 10.1093/nar/gky1281. Nucleic Acids Res. 2019. PMID: 30590800 Free PMC article.
-
yama, a mutant allele of Mov10l1, disrupts retrotransposon silencing and piRNA biogenesis.PLoS Genet. 2021 Feb 26;17(2):e1009265. doi: 10.1371/journal.pgen.1009265. eCollection 2021 Feb. PLoS Genet. 2021. PMID: 33635934 Free PMC article.
-
Switching of dominant retrotransposon silencing strategies from posttranscriptional to transcriptional mechanisms during male germ-cell development in mice.PLoS Genet. 2017 Jul 27;13(7):e1006926. doi: 10.1371/journal.pgen.1006926. eCollection 2017 Jul. PLoS Genet. 2017. PMID: 28749988 Free PMC article.
-
The regulatory functions of piRNA/PIWI in spermatogenesis.Yi Chuan. 2017 Aug 20;39(8):683-691. doi: 10.16288/j.yczz.17-245. Yi Chuan. 2017. PMID: 28903896 Review.
-
Noncanonical functions of PIWIL1/piRNAs in animal male germ cells and human diseases†.Biol Reprod. 2022 Jul 25;107(1):101-108. doi: 10.1093/biolre/ioac073. Biol Reprod. 2022. PMID: 35403682 Review.
Cited by
-
Small Noncoding RNAs in Reproduction and Infertility.Biomedicines. 2021 Dec 12;9(12):1884. doi: 10.3390/biomedicines9121884. Biomedicines. 2021. PMID: 34944700 Free PMC article. Review.
-
Host-transposon interactions: conflict, cooperation, and cooption.Genes Dev. 2019 Sep 1;33(17-18):1098-1116. doi: 10.1101/gad.327312.119. Genes Dev. 2019. PMID: 31481535 Free PMC article. Review.
-
Molecular and Functional Characterization of the Somatic PIWIL1/piRNA Pathway in Colorectal Cancer Cells.Cells. 2019 Nov 5;8(11):1390. doi: 10.3390/cells8111390. Cells. 2019. PMID: 31694219 Free PMC article.
-
Cracking the egg: A breakthrough in piRNA function in mammalian oocytes and embryos†.Biol Reprod. 2022 Jan 13;106(1):6-8. doi: 10.1093/biolre/ioab206. Biol Reprod. 2022. PMID: 34725680 Free PMC article. No abstract available.
-
PIWI-interacting RNAs: Mitochondria-based biogenesis and functions in cancer.Genes Dis. 2020 Oct 5;8(5):603-622. doi: 10.1016/j.gendis.2020.09.006. eCollection 2021 Sep. Genes Dis. 2020. PMID: 34291132 Free PMC article. Review.
References
-
- Kim V.N., Han J., Siomi M.C.. Biogenesis of small RNAs in animals. Nat. Rev. Mol. Cell Biol. 2009; 10:126–139. - PubMed
-
- Siomi M.C., Sato K., Pezic D., Aravin A.A.. PIWI-interacting small RNAs: the vanguard of genome defence. Nat. Rev. Mol. Cell Biol. 2011; 12:246–258. - PubMed
-
- Luteijn M.J., Ketting R.F.. PIWI-interacting RNAs: from generation to transgenerational epigenetics. Nat. Rev. Genet. 2013; 14:523–534. - PubMed
-
- Reuter M., Berninger P., Chuma S., Shah H., Hosokawa M., Funaya C., Antony C., Sachidanandam R., Pillai R.S.. Miwi catalysis is required for piRNA amplification-independent LINE1 transposon silencing. Nature. 2011; 480:264–267. - PubMed
-
- De Fazio S., Bartonicek N., Di Giacomo M., Abreu-Goodger C., Sankar A., Funaya C., Antony C., Moreira P.N., Enright A.J., O’Carroll D.. The endonuclease activity of Mili fuels piRNA amplification that silences LINE1 elements. Nature. 2011; 480:259–263. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
