

Glycan Alteration Imparts Cellular Resistance to a Membrane-Lytic Anticancer Peptide
- PMID: 28089756
- PMCID: PMC5316350
- DOI: 10.1016/j.chembiol.2016.12.009
Glycan Alteration Imparts Cellular Resistance to a Membrane-Lytic Anticancer Peptide
Abstract
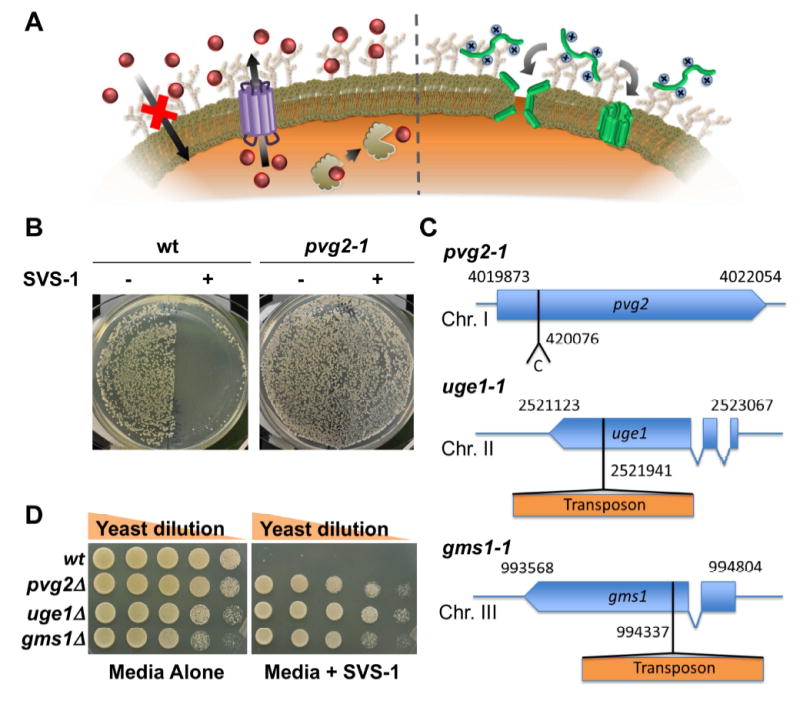
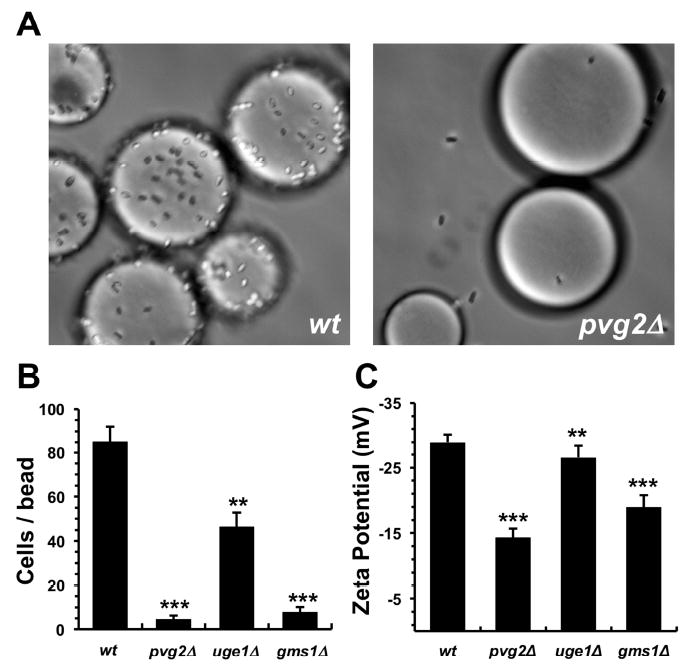
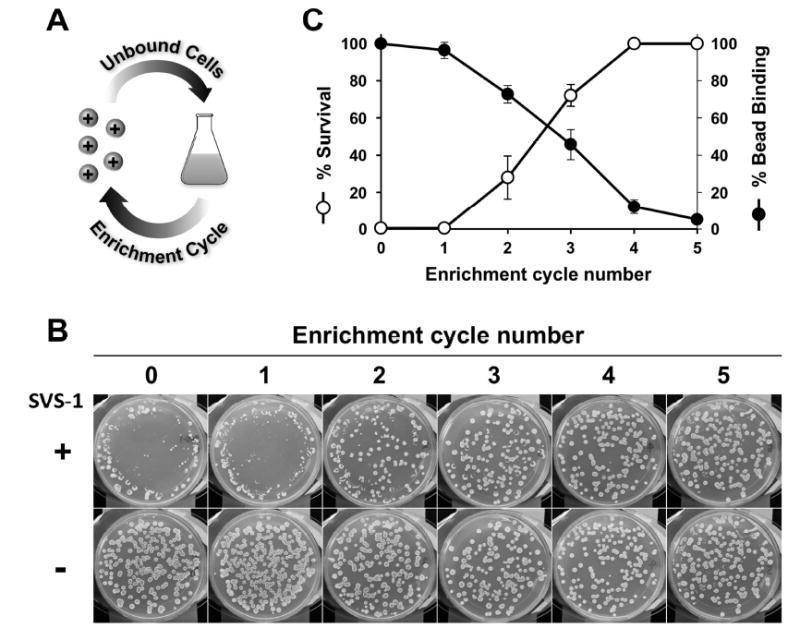
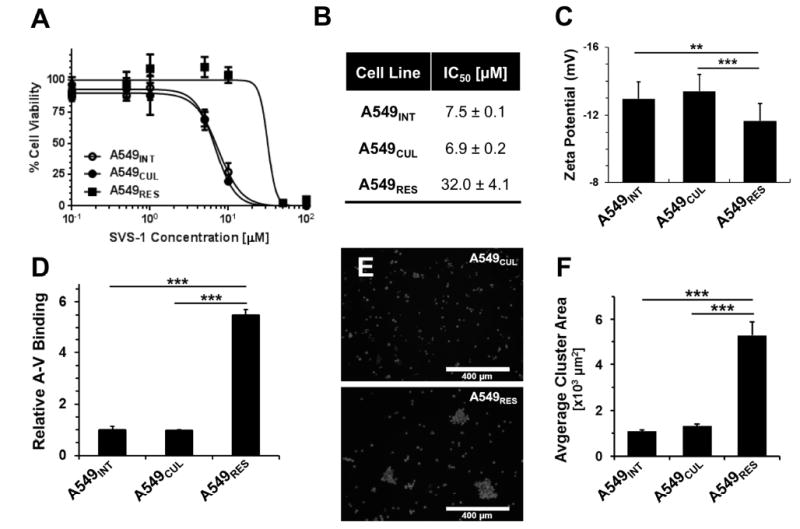
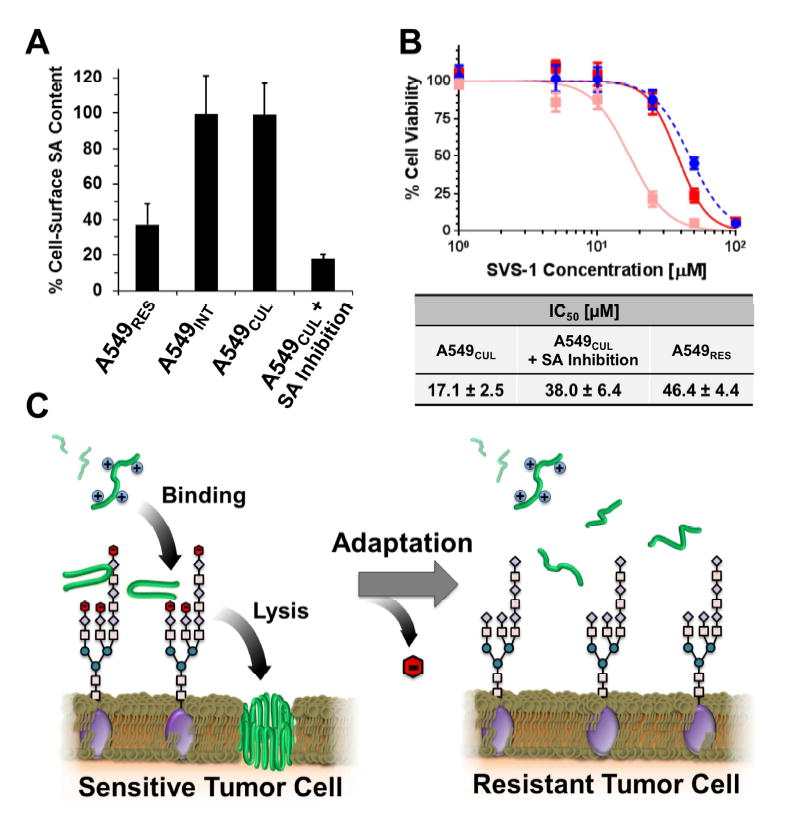
Although resistance toward small-molecule chemotherapeutics has been well studied, the potential of tumor cells to avoid destruction by membrane-lytic compounds remains unexplored. Anticancer peptides (ACPs) are a class of such agents that disrupt tumor cell membranes through rapid and non-stereospecific mechanisms, encouraging the perception that cellular resistance toward ACPs is unlikely to occur. We demonstrate that eukaryotic cells can, indeed, develop resistance to the model oncolytic peptide SVS-1, which preferentially disrupts the membranes of cancer cells. Utilizing fission yeast as a model organism, we show that ACP resistance is largely controlled through the loss of cell-surface anionic saccharides. A similar mechanism was discovered in mammalian cancer cells where removal of negatively charged sialic acid residues directly transformed SVS-1-sensitive cell lines into resistant phenotypes. These results demonstrate that changes in cell-surface glycosylation play a major role in tumor cell resistance toward oncolytic peptides.
Keywords: anticancer peptides; cancer cell resistance; genetics; glycosylation; membrane-lytic peptides.
Published by Elsevier Ltd.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Comment in
-
How Cancer Cells Become Resistant to Cationic Lytic Peptides: It's the Sugar!Cell Chem Biol. 2017 Feb 16;24(2):121-122. doi: 10.1016/j.chembiol.2017.01.009. Cell Chem Biol. 2017. PMID: 28212754
Similar articles
-
Lipopeptisomes: Anticancer peptide-assembled particles for fusolytic oncotherapy.Acta Biomater. 2018 Oct 15;80:269-277. doi: 10.1016/j.actbio.2018.09.025. Epub 2018 Sep 19. Acta Biomater. 2018. PMID: 30240951
-
Anticancer peptide NK-2 targets cell surface sulphated glycans rather than sialic acids.Biol Chem. 2012 Aug;393(8):817-27. doi: 10.1515/hsz-2012-0136. Biol Chem. 2012. PMID: 22944683
-
Engineering Anticancer Amphipathic Peptide-Dendronized Compounds for Highly-Efficient Plasma/Organelle Membrane Perturbation and Multidrug Resistance Reversal.ACS Appl Mater Interfaces. 2018 Sep 19;10(37):30952-30962. doi: 10.1021/acsami.8b07917. Epub 2018 Aug 22. ACS Appl Mater Interfaces. 2018. PMID: 30088909
-
On the selectivity and efficacy of defense peptides with respect to cancer cells.Med Res Rev. 2013 Jan;33(1):190-234. doi: 10.1002/med.20252. Epub 2011 Sep 15. Med Res Rev. 2013. PMID: 21922503 Review.
-
Anticancer alpha-helical peptides and structure/function relationships underpinning their interactions with tumour cell membranes.Curr Protein Pept Sci. 2006 Dec;7(6):487-99. doi: 10.2174/138920306779025611. Curr Protein Pept Sci. 2006. PMID: 17168782 Review.
Cited by
-
The effect of turn residues on the folding and cell-penetrating activity of β-hairpin peptides and applications toward protein delivery.Pept Sci (Hoboken). 2020 Jan;112(1):e24125. doi: 10.1002/pep2.24125. Epub 2019 Jun 12. Pept Sci (Hoboken). 2020. PMID: 34504991 Free PMC article.
-
Synthesis and Characterization of Radiogallium-Labeled Cationic Amphiphilic Peptides as Tumor Imaging Agents.Cancers (Basel). 2021 May 14;13(10):2388. doi: 10.3390/cancers13102388. Cancers (Basel). 2021. PMID: 34069243 Free PMC article.
-
Are peptides a solution for the treatment of hyperactivated JAK3 pathways?Inflammopharmacology. 2019 Jun;27(3):433-452. doi: 10.1007/s10787-019-00589-2. Epub 2019 Mar 30. Inflammopharmacology. 2019. PMID: 30929155 Review.
-
Integrated Design of a Membrane-Lytic Peptide-Based Intravenous Nanotherapeutic Suppresses Triple-Negative Breast Cancer.Adv Sci (Weinh). 2022 May;9(13):e2105506. doi: 10.1002/advs.202105506. Epub 2022 Mar 4. Adv Sci (Weinh). 2022. PMID: 35246961 Free PMC article.
-
UV light-induced spatial loss of sialic acid capping using a photoactivatable sialyltransferase inhibitor.RSC Chem Biol. 2023 May 25;4(7):506-511. doi: 10.1039/d3cb00006k. eCollection 2023 Jul 5. RSC Chem Biol. 2023. PMID: 37415865 Free PMC article.
References
-
- Andreishcheva EN, Kunkel JP, Gemmill TR, Trimble RB. Five genes involved in biosynthesis of the pyruvylated Galbeta1,3-epitope in Schizosaccharomyces pombe N-linked glycans. J Biol Chem. 2004;279:35644–35655. - PubMed
-
- Bacheler L, Jeffrey S, Hanna G, D’Aquila R, Wallace L, Logue K, Cordova B, Hertogs K, Larder B, Buckery R. Genotypic correlates of phenotypic resistance to efavirenz in virus isolates from patients failing nonnucleoside reverse transcriptase inhibitor therapy. J Virol. 2001;75:4999–5008. - PMC - PubMed
-
- Bahler J, Wu JQ, Longtine MS, Shah NG, McKenzie A, 3rd, Steever AB, Wach A, Philippsen P, Pringle JR. Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast. 1998;14:943–951. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
