

The Prion-Like Behavior of Assembled Tau in Transgenic Mice
- PMID: 27940600
- PMCID: PMC5629990
- DOI: 10.1101/cshperspect.a024372
The Prion-Like Behavior of Assembled Tau in Transgenic Mice
Abstract
Tauopathies constitute neurodegenerative diseases that are characterized by the intracellular deposition of filaments made of hyperphosphorylated tau protein. The pattern of tau deposition in Alzheimer's disease follows a stereotypical progression, with the first lesions appearing in the locus coeruleus and entorhinal cortex, from where they appear to spread to the hippocampus and neocortex. Propagation of pathological tau is also characteristic of argyrophilic grain disease, where the lesions seem to spread through distinct regions of the limbic system. In chronic traumatic encephalopathy, tauopathy appears to spread from the neocortex to the brainstem. These findings implicate neuron-to-neuron propagation of tau aggregates. Isoform compositions and morphologies of tau filaments can differ between tauopathies, which is consistent with the existence of distinct tau strains. Here, we review recent findings that support prion-like mechanisms in the pathogenesis of tauopathies through the experimental use of transgenic mice.
Copyright © 2017 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
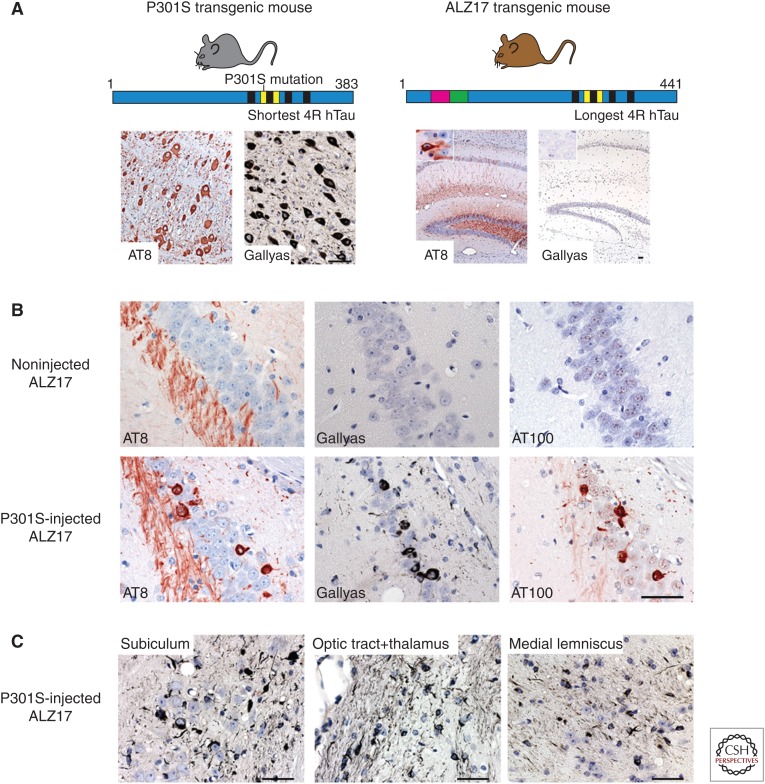
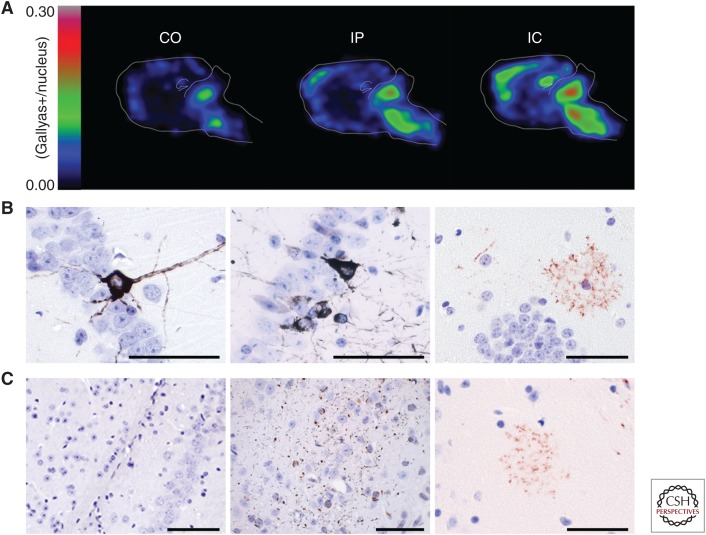
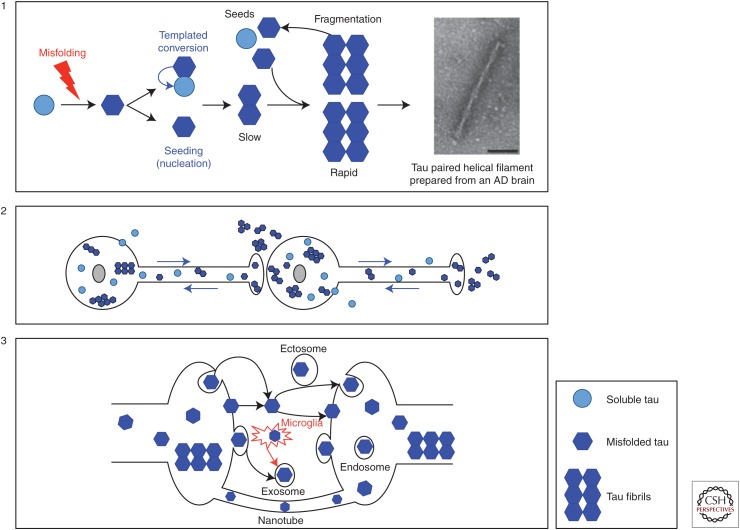
Similar articles
-
Invited review: Prion-like transmission and spreading of tau pathology.Neuropathol Appl Neurobiol. 2015 Feb;41(1):47-58. doi: 10.1111/nan.12197. Neuropathol Appl Neurobiol. 2015. PMID: 25399729 Review.
-
"Prion-like" templated misfolding in tauopathies.Brain Pathol. 2013 May;23(3):342-9. doi: 10.1111/bpa.12044. Brain Pathol. 2013. PMID: 23587140 Free PMC article. Review.
-
Tau pathology spread in PS19 tau transgenic mice following locus coeruleus (LC) injections of synthetic tau fibrils is determined by the LC's afferent and efferent connections.Acta Neuropathol. 2015 Sep;130(3):349-62. doi: 10.1007/s00401-015-1458-4. Epub 2015 Jul 7. Acta Neuropathol. 2015. PMID: 26150341 Free PMC article.
-
Towards a Mechanistic Model of Tau-Mediated Pathology in Tauopathies: What Can We Learn from Cell-Based In Vitro Assays?Int J Mol Sci. 2022 Sep 29;23(19):11527. doi: 10.3390/ijms231911527. Int J Mol Sci. 2022. PMID: 36232835 Free PMC article. Review.
-
The twenty-four KDa C-terminal tau fragment increases with aging in tauopathy mice: implications of prion-like properties.Hum Mol Genet. 2015 Nov 15;24(22):6403-16. doi: 10.1093/hmg/ddv351. Epub 2015 Sep 15. Hum Mol Genet. 2015. PMID: 26374846
Cited by
-
Cis P-tau underlies vascular contribution to cognitive impairment and dementia and can be effectively targeted by immunotherapy in mice.Sci Transl Med. 2021 Jun 2;13(596):eaaz7615. doi: 10.1126/scitranslmed.aaz7615. Sci Transl Med. 2021. PMID: 34078745 Free PMC article.
-
Prion-Like Propagation of Post-Translationally Modified Tau in Alzheimer's Disease: A Hypothesis.J Mol Neurosci. 2018 Aug;65(4):480-490. doi: 10.1007/s12031-018-1111-5. Epub 2018 Jul 7. J Mol Neurosci. 2018. PMID: 29982964 Review.
-
Therapeutic Strategies for Restoring Tau Homeostasis.Cold Spring Harb Perspect Med. 2018 Jan 2;8(1):a024612. doi: 10.1101/cshperspect.a024612. Cold Spring Harb Perspect Med. 2018. PMID: 28159830 Free PMC article. Review.
-
Cellular Models for the Study of Prions.Cold Spring Harb Perspect Med. 2017 Feb 1;7(2):a024026. doi: 10.1101/cshperspect.a024026. Cold Spring Harb Perspect Med. 2017. PMID: 27815306 Free PMC article. Review.
-
Amyloid-beta and tau protein beyond Alzheimer's disease.Neural Regen Res. 2024 Jun 1;19(6):1262-1276. doi: 10.4103/1673-5374.386406. Epub 2023 Oct 2. Neural Regen Res. 2024. PMID: 37905874 Free PMC article.
References
-
- Ahmed Z, Cooper J, Murray TK, Garn K, McNaughton E, Clarke H, Parhizkar S, Ward MA, Cavallini A, Jackson S, et al. 2014. A novel in vivo model of tau propagation with rapid and progressive neurofibrillary tangle pathology: The pattern of spread is determined by connectivity, not proximity. Acta Neuropathol 127: 667–683. - PMC - PubMed
-
- Boluda S, Iba M, Zhang B, Raible KM, Lee VMY, Trojanowski JQ. 2015. Differential induction and spread of tau pathology in young PS19 tau transgenic mice following intracerebral injections of pathological tau from Alzheimer’s disease or corticobasal degeneration brains. Acta Neuropathol 129: 221–237. - PMC - PubMed
-
- Botez G, Probst A, Ipsen S, Tolnay M. 1999. Astrocytes expressing hyperphosphorylated tau protein without glial fibrillary tangles in argyrophilic grain disease. Acta Neuropathol 98: 251–256. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources