

Whole-Organism Developmental Expression Profiling Identifies RAB-28 as a Novel Ciliary GTPase Associated with the BBSome and Intraflagellar Transport
- PMID: 27930654
- PMCID: PMC5145144
- DOI: 10.1371/journal.pgen.1006469
Whole-Organism Developmental Expression Profiling Identifies RAB-28 as a Novel Ciliary GTPase Associated with the BBSome and Intraflagellar Transport
Abstract
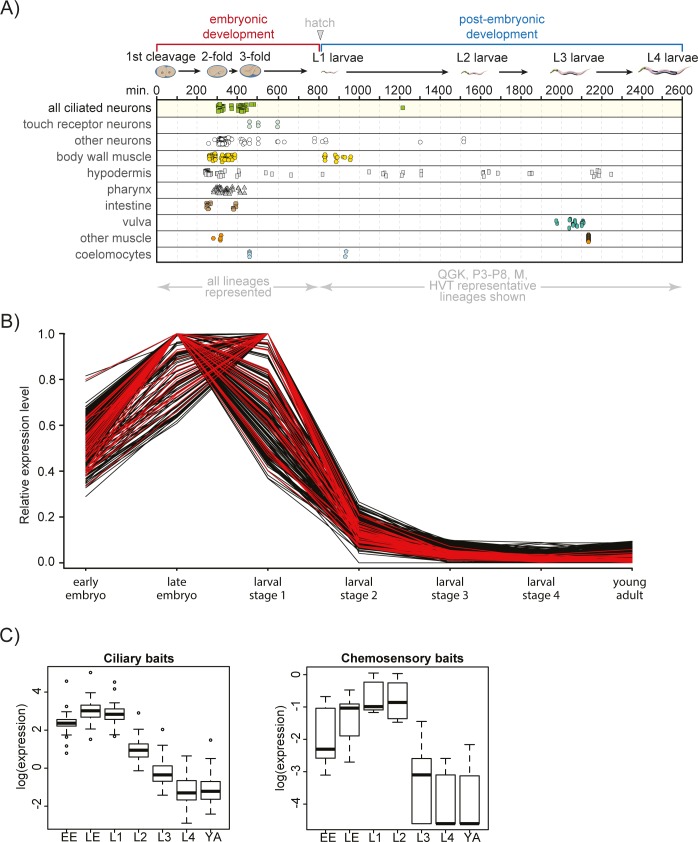
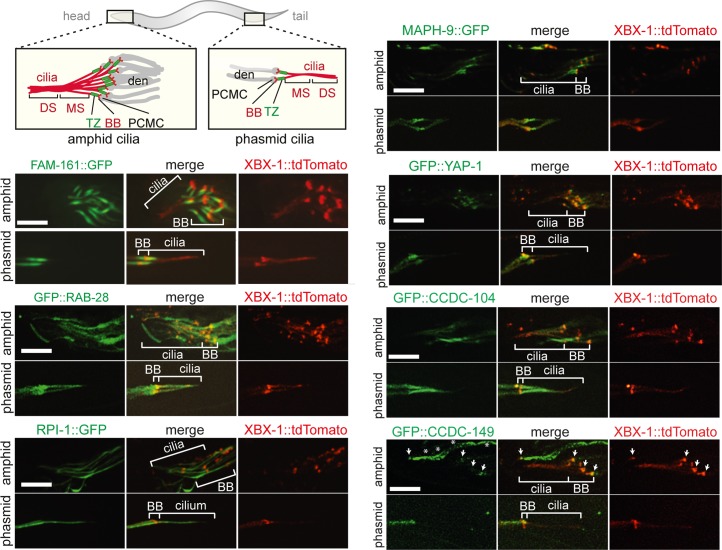
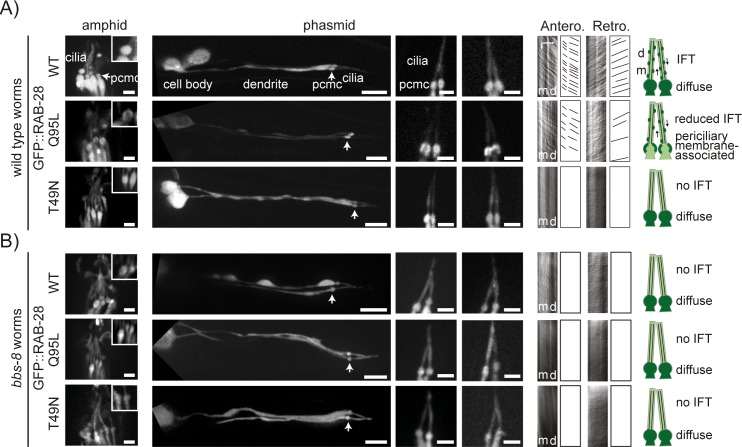
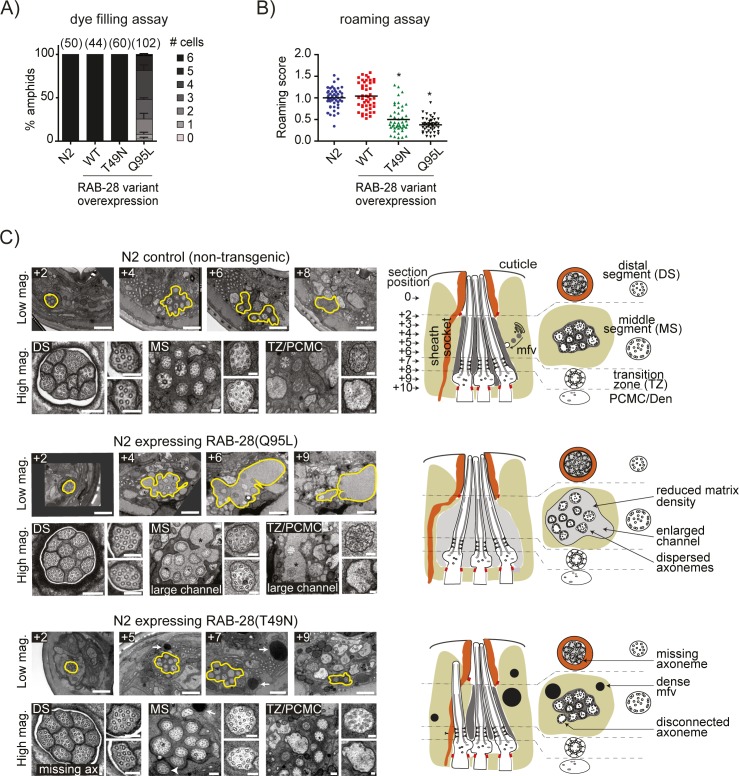
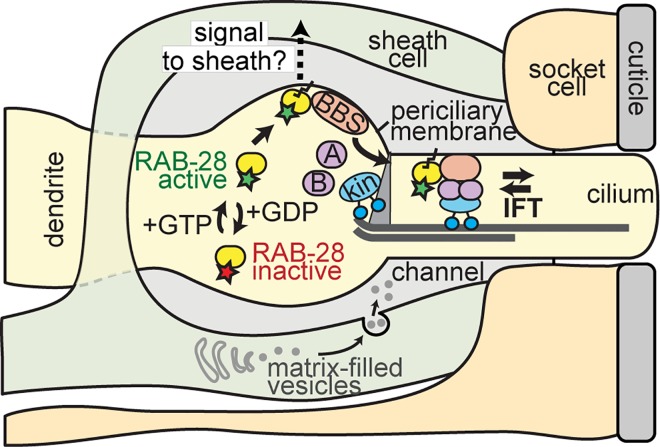
Primary cilia are specialised sensory and developmental signalling devices extending from the surface of most eukaryotic cells. Defects in these organelles cause inherited human disorders (ciliopathies) such as retinitis pigmentosa and Bardet-Biedl syndrome (BBS), frequently affecting many physiological and developmental processes across multiple organs. Cilium formation, maintenance and function depend on intracellular transport systems such as intraflagellar transport (IFT), which is driven by kinesin-2 and IFT-dynein motors and regulated by the Bardet-Biedl syndrome (BBS) cargo-adaptor protein complex, or BBSome. To identify new cilium-associated genes, we employed the nematode C. elegans, where ciliogenesis occurs within a short timespan during late embryogenesis when most sensory neurons differentiate. Using whole-organism RNA-Seq libraries, we discovered a signature expression profile highly enriched for transcripts of known ciliary proteins, including FAM-161 (FAM161A orthologue), CCDC-104 (CCDC104), and RPI-1 (RP1/RP1L1), which we confirm are cilium-localised in worms. From a list of 185 candidate ciliary genes, we uncover orthologues of human MAP9, YAP, CCDC149, and RAB28 as conserved cilium-associated components. Further analyses of C. elegans RAB-28, recently associated with autosomal-recessive cone-rod dystrophy, reveal that this small GTPase is exclusively expressed in ciliated neurons where it dynamically associates with IFT trains. Whereas inactive GDP-bound RAB-28 displays no IFT movement and diffuse localisation, GTP-bound (activated) RAB-28 concentrates at the periciliary membrane in a BBSome-dependent manner and undergoes bidirectional IFT. Functional analyses reveal that whilst cilium structure, sensory function and IFT are seemingly normal in a rab-28 null allele, overexpression of predicted GDP or GTP locked variants of RAB-28 perturbs cilium and sensory pore morphogenesis and function. Collectively, our findings present a new approach for identifying ciliary proteins, and unveil RAB28, a GTPase most closely related to the BBS protein RABL4/IFT27, as an IFT-associated cargo with BBSome-dependent cell autonomous and non-autonomous functions at the ciliary base.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

Similar articles
-
Intraflagellar transport protein RABL5/IFT22 recruits the BBSome to the basal body through the GTPase ARL6/BBS3.Proc Natl Acad Sci U S A. 2020 Feb 4;117(5):2496-2505. doi: 10.1073/pnas.1901665117. Epub 2020 Jan 17. Proc Natl Acad Sci U S A. 2020. PMID: 31953262 Free PMC article.
-
Ciliary protein trafficking mediated by IFT and BBSome complexes with the aid of kinesin-2 and dynein-2 motors.J Biochem. 2018 Mar 1;163(3):155-164. doi: 10.1093/jb/mvx087. J Biochem. 2018. PMID: 29272450 Review.
-
Active transport and diffusion barriers restrict Joubert Syndrome-associated ARL13B/ARL-13 to an Inv-like ciliary membrane subdomain.PLoS Genet. 2013;9(12):e1003977. doi: 10.1371/journal.pgen.1003977. Epub 2013 Dec 5. PLoS Genet. 2013. PMID: 24339792 Free PMC article.
-
The BBSome controls IFT assembly and turnaround in cilia.Nat Cell Biol. 2012 Sep;14(9):950-7. doi: 10.1038/ncb2560. Epub 2012 Aug 26. Nat Cell Biol. 2012. PMID: 22922713 Free PMC article.
-
Trafficking of ciliary membrane proteins by the intraflagellar transport/BBSome machinery.Essays Biochem. 2018 Dec 7;62(6):753-763. doi: 10.1042/EBC20180030. Print 2018 Dec 7. Essays Biochem. 2018. PMID: 30287585 Free PMC article. Review.
Cited by
-
Coiled-Coil Domain-Containing (CCDC) Proteins: Functional Roles in General and Male Reproductive Physiology.Reprod Sci. 2021 Oct;28(10):2725-2734. doi: 10.1007/s43032-021-00595-2. Epub 2021 May 3. Reprod Sci. 2021. PMID: 33942254 Review.
-
Systematic proteomic analysis of LRRK2-mediated Rab GTPase phosphorylation establishes a connection to ciliogenesis.Elife. 2017 Nov 10;6:e31012. doi: 10.7554/eLife.31012. Elife. 2017. PMID: 29125462 Free PMC article.
-
The tubulin code specializes neuronal cilia for extracellular vesicle release.Dev Neurobiol. 2021 Apr;81(3):231-252. doi: 10.1002/dneu.22787. Epub 2020 Nov 8. Dev Neurobiol. 2021. PMID: 33068333 Free PMC article. Review.
-
Genetic Deletion of Zebrafish Rab28 Causes Defective Outer Segment Shedding, but Not Retinal Degeneration.Front Cell Dev Biol. 2020 Mar 17;8:136. doi: 10.3389/fcell.2020.00136. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 32258030 Free PMC article.
-
Consequences of Rab GTPase dysfunction in genetic or acquired human diseases.Small GTPases. 2018 Mar 4;9(1-2):158-181. doi: 10.1080/21541248.2017.1397833. Epub 2017 Dec 28. Small GTPases. 2018. PMID: 29239692 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
