

Multidomain structure and correlated dynamics determined by self-consistent FRET networks
- PMID: 27918541
- PMCID: PMC5289555
- DOI: 10.1038/nmeth.4081
Multidomain structure and correlated dynamics determined by self-consistent FRET networks
Abstract
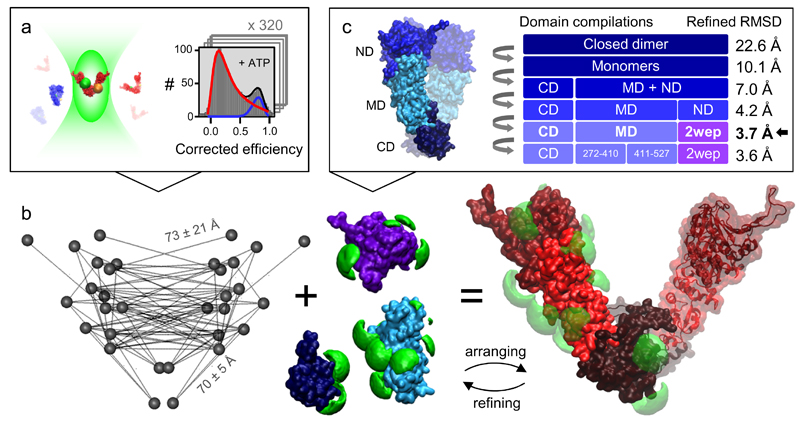
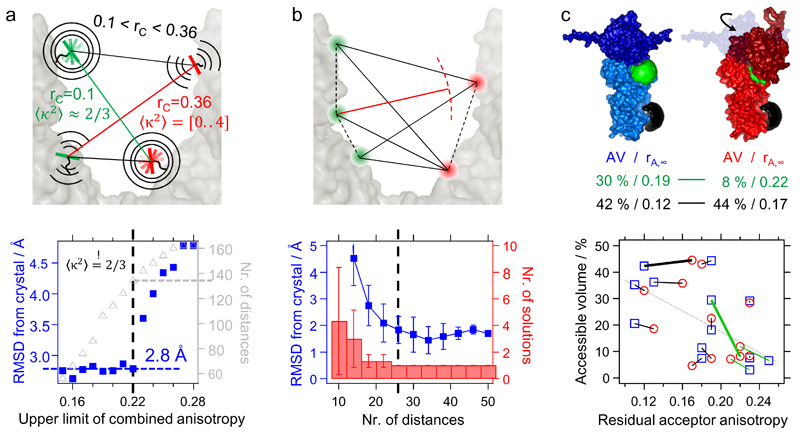
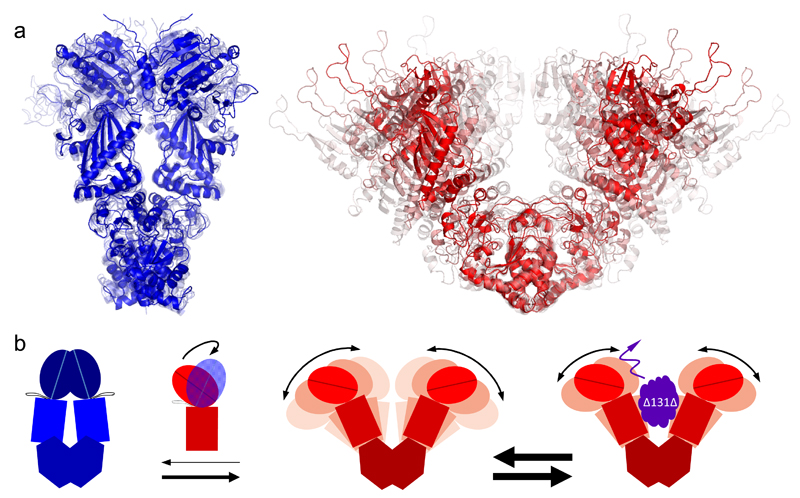
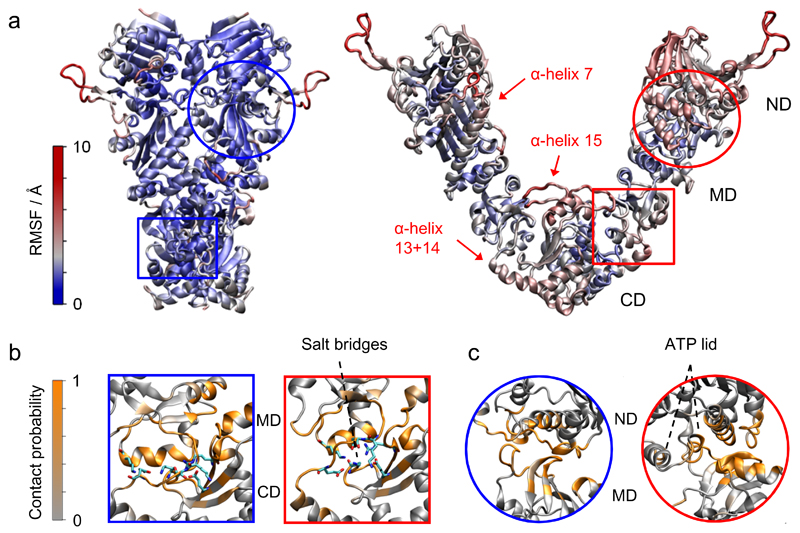
We present an approach that enables us to simultaneously access structure and dynamics of a multidomain protein in solution. Dynamic domain arrangements are experimentally determined by combining self-consistent networks of distance distributions with known domain structures. Local structural dynamics are correlated with the global arrangements by analyzing networks of time-resolved single-molecule fluorescence parameters. The strength of this hybrid approach is shown by an application to the flexible multidomain protein Hsp90. The average solution structure of Hsp90's closed state resembles the known X-ray crystal structure with Angstrom precision. The open state is represented by an ensemble of conformations with interdomain fluctuations of up to 25 Å. The data reveal a state-specific suppression of the submillisecond fluctuations by dynamic protein-protein interaction. Finally, the method enables localization and functional characterization of dynamic elements and domain interfaces.
Conflict of interest statement
The authors declare no competing financial interests.
Figures

Comment in
-
Cool and dynamic: single-molecule fluorescence-based structural biology.Nat Methods. 2017 Jan 31;14(2):123-124. doi: 10.1038/nmeth.4159. Nat Methods. 2017. PMID: 28139670 No abstract available.
Similar articles
-
The charged linker of the molecular chaperone Hsp90 modulates domain contacts and biological function.Proc Natl Acad Sci U S A. 2014 Dec 16;111(50):17881-6. doi: 10.1073/pnas.1414073111. Epub 2014 Dec 2. Proc Natl Acad Sci U S A. 2014. PMID: 25468961 Free PMC article.
-
From a ratchet mechanism to random fluctuations evolution of Hsp90's mechanochemical cycle.J Mol Biol. 2012 Oct 26;423(3):462-71. doi: 10.1016/j.jmb.2012.07.026. Epub 2012 Aug 6. J Mol Biol. 2012. PMID: 22878379
-
Two closed ATP- and ADP-dependent conformations in yeast Hsp90 chaperone detected by Mn(II) EPR spectroscopic techniques.Proc Natl Acad Sci U S A. 2020 Jan 7;117(1):395-404. doi: 10.1073/pnas.1916030116. Epub 2019 Dec 20. Proc Natl Acad Sci U S A. 2020. PMID: 31862713 Free PMC article.
-
Hsp90 structure and function studied by NMR spectroscopy.Biochim Biophys Acta. 2012 Mar;1823(3):636-47. doi: 10.1016/j.bbamcr.2011.11.009. Epub 2011 Dec 6. Biochim Biophys Acta. 2012. PMID: 22155720 Review.
-
Hsp90's secrets unfold: new insights from structural and functional studies.Trends Cell Biol. 1999 Jul;9(7):262-8. doi: 10.1016/s0962-8924(99)01580-9. Trends Cell Biol. 1999. PMID: 10370241 Review.
Cited by
-
Can DyeCycling break the photobleaching limit in single-molecule FRET?Nano Res. 2022;15(11):9818-9830. doi: 10.1007/s12274-022-4420-5. Epub 2022 May 13. Nano Res. 2022. PMID: 35582137 Free PMC article.
-
Experimental Protein Molecular Dynamics: Broadband Dielectric Spectroscopy coupled with nanoconfinement.Sci Rep. 2019 Nov 29;9(1):17988. doi: 10.1038/s41598-019-54562-8. Sci Rep. 2019. PMID: 31784681 Free PMC article.
-
Accurate Transfer Efficiencies, Distance Distributions, and Ensembles of Unfolded and Intrinsically Disordered Proteins From Single-Molecule FRET.Methods Enzymol. 2018;611:287-325. doi: 10.1016/bs.mie.2018.09.030. Epub 2018 Nov 16. Methods Enzymol. 2018. PMID: 30471690 Free PMC article.
-
Unraveling multi-state molecular dynamics in single-molecule FRET experiments. I. Theory of FRET-lines.J Chem Phys. 2022 Apr 14;156(14):141501. doi: 10.1063/5.0089134. J Chem Phys. 2022. PMID: 35428384 Free PMC article.
-
High-Resolution Single-Molecule FRET via DNA eXchange (FRET X).Nano Lett. 2021 Apr 14;21(7):3295-3301. doi: 10.1021/acs.nanolett.1c00725. Epub 2021 Mar 19. Nano Lett. 2021. PMID: 33739111 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
