

Whole-Genome Sequencing of Cytogenetically Balanced Chromosome Translocations Identifies Potentially Pathological Gene Disruptions and Highlights the Importance of Microhomology in the Mechanism of Formation
- PMID: 27862604
- PMCID: PMC5225243
- DOI: 10.1002/humu.23146
Whole-Genome Sequencing of Cytogenetically Balanced Chromosome Translocations Identifies Potentially Pathological Gene Disruptions and Highlights the Importance of Microhomology in the Mechanism of Formation
Abstract
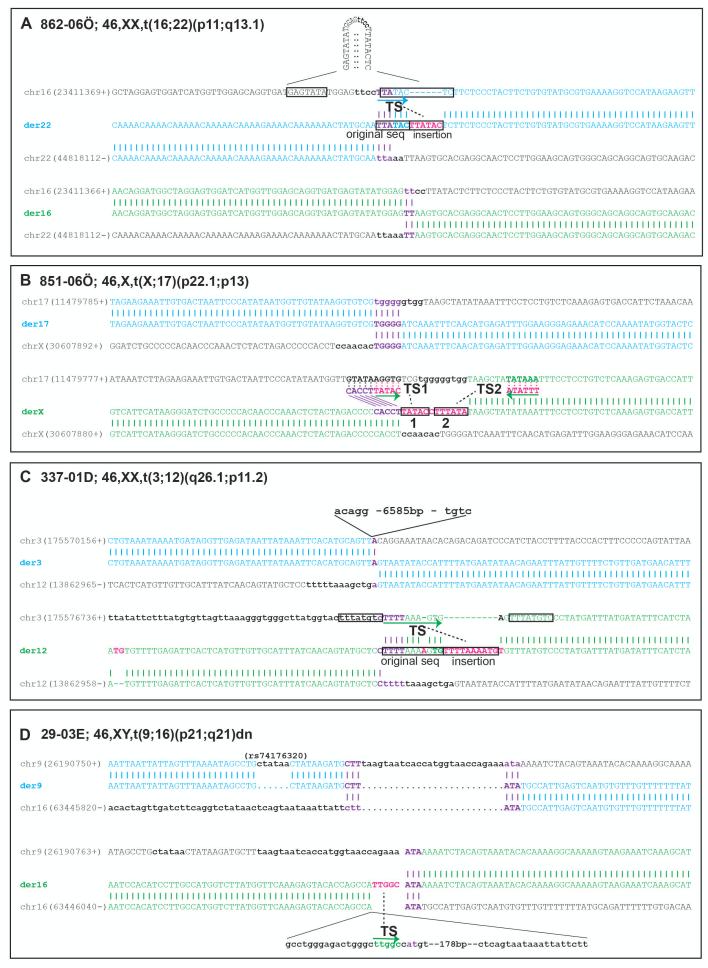
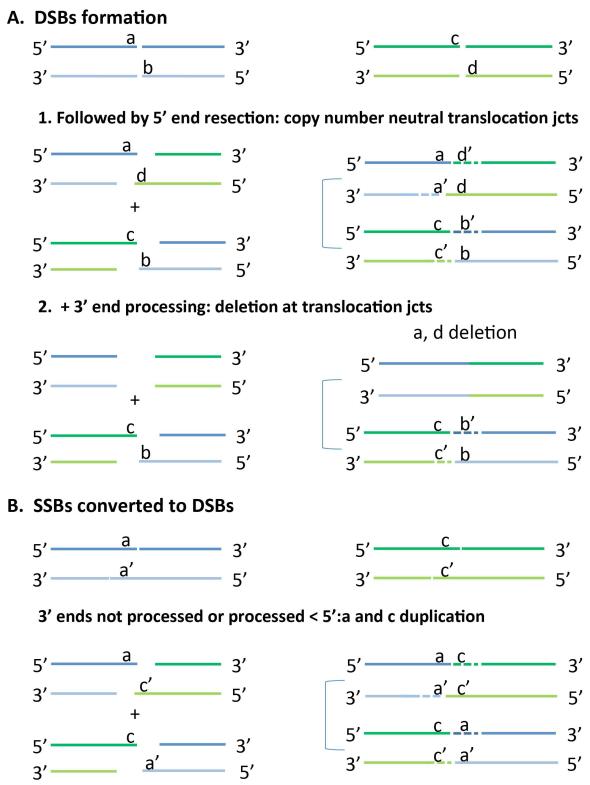
Most balanced translocations are thought to result mechanistically from nonhomologous end joining or, in rare cases of recurrent events, by nonallelic homologous recombination. Here, we use low-coverage mate pair whole-genome sequencing to fine map rearrangement breakpoint junctions in both phenotypically normal and affected translocation carriers. In total, 46 junctions from 22 carriers of balanced translocations were characterized. Genes were disrupted in 48% of the breakpoints; recessive genes in four normal carriers and known dominant intellectual disability genes in three affected carriers. Finally, seven candidate disease genes were disrupted in five carriers with neurocognitive disabilities (SVOPL, SUSD1, TOX, NCALD, SLC4A10) and one XX-male carrier with Tourette syndrome (LYPD6, GPC5). Breakpoint junction analyses revealed microhomology and small templated insertions in a substantive fraction of the analyzed translocations (17.4%; n = 4); an observation that was substantiated by reanalysis of 37 previously published translocation junctions. Microhomology associated with templated insertions is a characteristic seen in the breakpoint junctions of rearrangements mediated by error-prone replication-based repair mechanisms. Our data implicate that a mechanism involving template switching might contribute to the formation of at least 15% of the interchromosomal translocation events.
Keywords: balanced chromosomal aberration; microhomology; nonhomologous end joining; reciprocal translocation; replication-based repair mechanisms; whole-genome sequencing.
© 2016 WILEY PERIODICALS, INC.
Figures


Similar articles
-
Unbalanced translocations arise from diverse mutational mechanisms including chromothripsis.Genome Res. 2015 Jul;25(7):937-47. doi: 10.1101/gr.191247.115. Epub 2015 Jun 12. Genome Res. 2015. PMID: 26070663 Free PMC article.
-
Observation and prediction of recurrent human translocations mediated by NAHR between nonhomologous chromosomes.Genome Res. 2011 Jan;21(1):33-46. doi: 10.1101/gr.111609.110. Genome Res. 2011. PMID: 21205869 Free PMC article.
-
Can a Liquid Biopsy Detect Circulating Tumor DNA With Low-passage Whole-genome Sequencing in Patients With a Sarcoma? A Pilot Evaluation.Clin Orthop Relat Res. 2025 Jan 1;483(1):39-48. doi: 10.1097/CORR.0000000000003161. Epub 2024 Jun 21. Clin Orthop Relat Res. 2025. PMID: 38905450
-
Depressing time: Waiting, melancholia, and the psychoanalytic practice of care.In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. PMID: 36137063 Free Books & Documents. Review.
-
Immunomodulators and immunosuppressants for multiple sclerosis: a network meta-analysis.Cochrane Database Syst Rev. 2013 Jun 6;2013(6):CD008933. doi: 10.1002/14651858.CD008933.pub2. Cochrane Database Syst Rev. 2013. PMID: 23744561 Free PMC article. Review.
Cited by
-
From cytogenetics to cytogenomics: whole-genome sequencing as a first-line test comprehensively captures the diverse spectrum of disease-causing genetic variation underlying intellectual disability.Genome Med. 2019 Nov 7;11(1):68. doi: 10.1186/s13073-019-0675-1. Genome Med. 2019. PMID: 31694722 Free PMC article.
-
Whole-genome mate-pair sequencing of apparently balanced chromosome rearrangements reveals complex structural variations: two case studies.Mol Cytogenet. 2020 May 6;13:15. doi: 10.1186/s13039-020-00487-1. eCollection 2020. Mol Cytogenet. 2020. PMID: 32391085 Free PMC article.
-
Resolving complex structural variants via nanopore sequencing.Front Genet. 2023 Aug 16;14:1213917. doi: 10.3389/fgene.2023.1213917. eCollection 2023. Front Genet. 2023. PMID: 37674481 Free PMC article. Review.
-
Induction of Chromosomal Translocations with CRISPR-Cas9 and Other Nucleases: Understanding the Repair Mechanisms That Give Rise to Translocations.Adv Exp Med Biol. 2018;1044:15-25. doi: 10.1007/978-981-13-0593-1_2. Adv Exp Med Biol. 2018. PMID: 29956288 Free PMC article. Review.
-
High-resolution detection of chromosomal rearrangements in leukemias through mate pair whole genome sequencing.PLoS One. 2018 Mar 12;13(3):e0193928. doi: 10.1371/journal.pone.0193928. eCollection 2018. PLoS One. 2018. PMID: 29529047 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
