

Constitutive Glycolytic Metabolism Supports CD8+ T Cell Effector Memory Differentiation during Viral Infection
- PMID: 27836431
- PMCID: PMC5130099
- DOI: 10.1016/j.immuni.2016.10.017
Constitutive Glycolytic Metabolism Supports CD8+ T Cell Effector Memory Differentiation during Viral Infection
Abstract
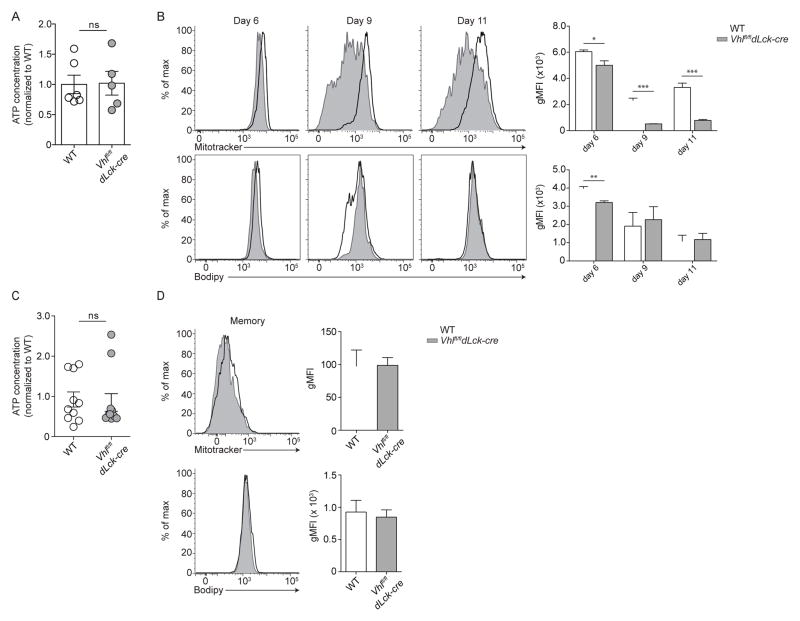
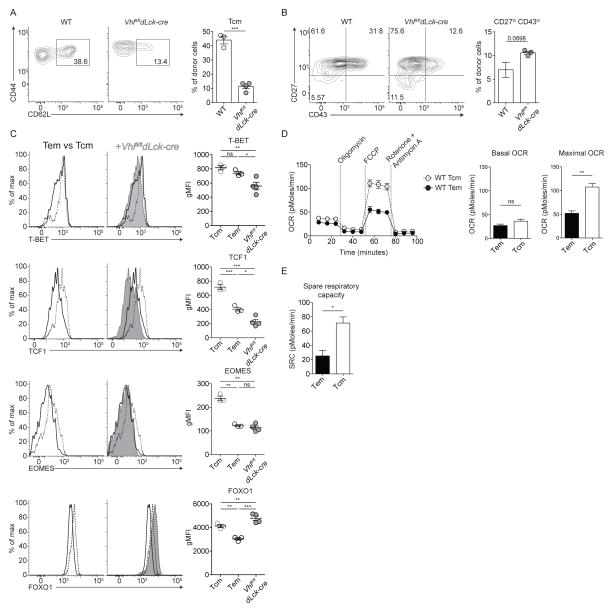
Extensive metabolic changes accompany T cell activation, including a switch to glycolytic energy production and increased biosynthesis. Recent studies suggest that subsequent return to reliance on oxidative phosphorylation and increasing spare respiratory capacity are essential for the differentiation of memory CD8+ T cells. In contrast, we found that constitutive glycolytic metabolism and suppression of oxidative phosphorylation in CD8+ T cells, achieved by conditional deletion of hypoxia-inducible factor regulator Vhl, accelerated CD8+ memory cell differentiation during viral infection. Despite sustained glycolysis, CD8+ memory cells emerged that upregulated key memory-associated cytokine receptors and transcription factors and showed a heightened response to secondary challenge. In addition, increased glycolysis not only permitted memory formation, but it also favored the formation of long-lived effector-memory CD8+ T cells. These data redefine the role of cellular metabolism in memory cell differentiation, showing that reliance on glycolytic metabolism does not hinder formation of a protective memory population.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
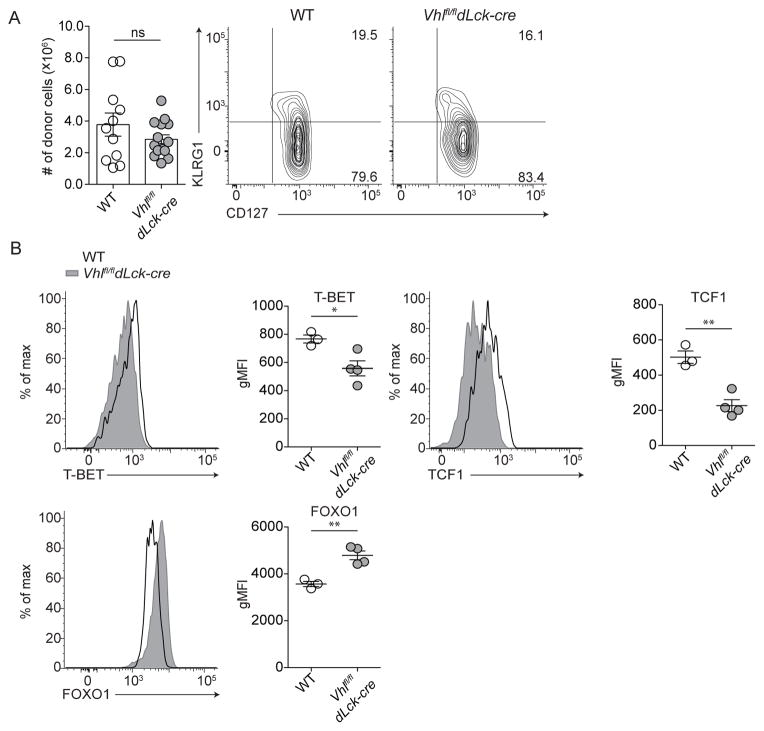
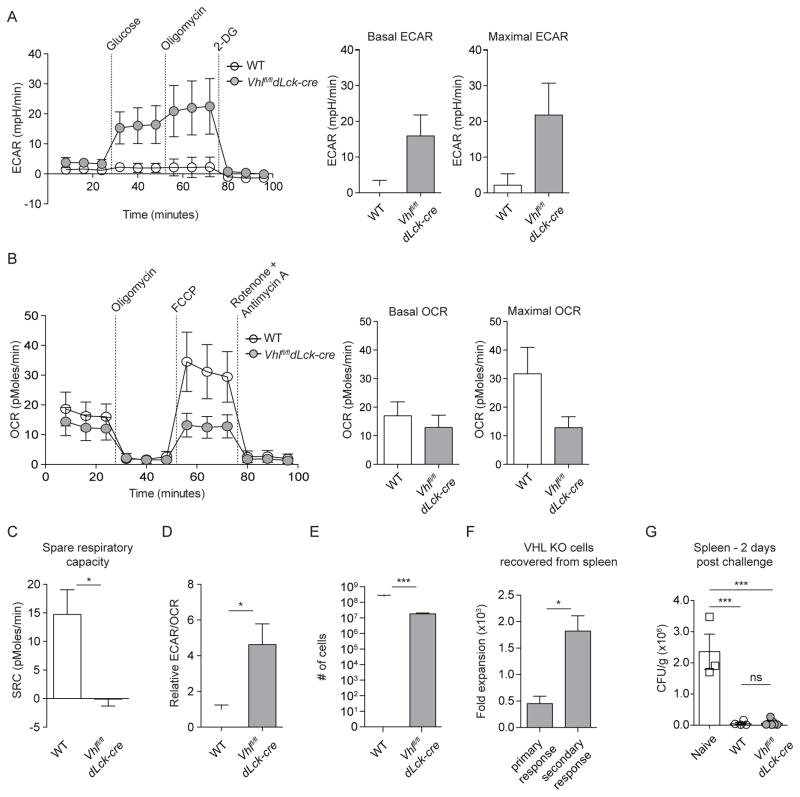
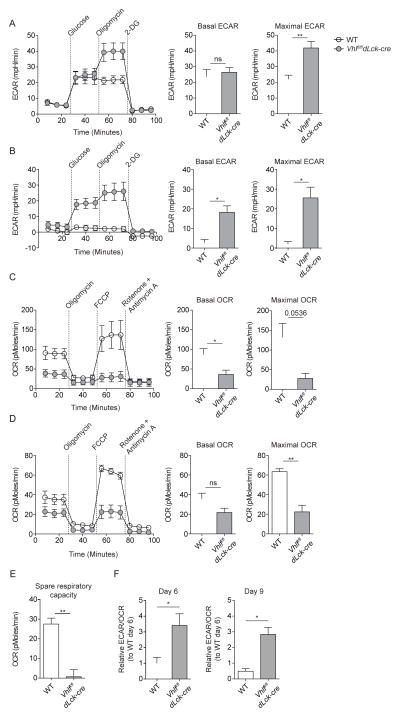
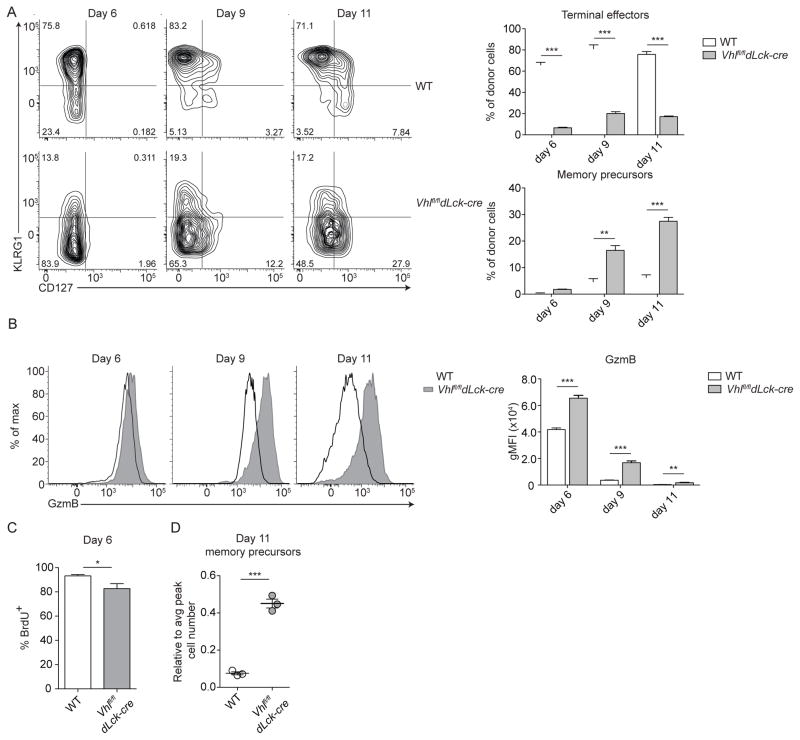
Comment in
-
VHL Brings Warburg into the Memory Spotlight.Immunity. 2016 Nov 15;45(5):953-955. doi: 10.1016/j.immuni.2016.11.001. Immunity. 2016. PMID: 27851921
Similar articles
-
Signal 3 requirement for memory CD8+ T-cell activation is determined by the infectious pathogen.Eur J Immunol. 2011 Nov;41(11):3176-86. doi: 10.1002/eji.201141537. Epub 2011 Sep 26. Eur J Immunol. 2011. PMID: 21830209
-
Transient expression of ZBTB32 in anti-viral CD8+ T cells limits the magnitude of the effector response and the generation of memory.PLoS Pathog. 2017 Aug 21;13(8):e1006544. doi: 10.1371/journal.ppat.1006544. eCollection 2017 Aug. PLoS Pathog. 2017. PMID: 28827827 Free PMC article.
-
Distinct roles for IL-2 and IL-15 in the differentiation and survival of CD8+ effector and memory T cells.J Immunol. 2010 Jun 15;184(12):6719-30. doi: 10.4049/jimmunol.0904089. Epub 2010 May 14. J Immunol. 2010. PMID: 20483725 Free PMC article.
-
Generation of effector CD8+ T cells and their conversion to memory T cells.Immunol Rev. 2010 Jul;236:151-66. doi: 10.1111/j.1600-065X.2010.00926.x. Immunol Rev. 2010. PMID: 20636815 Free PMC article. Review.
-
Immunometabolism of infections.Nat Rev Immunol. 2020 Feb;20(2):79-80. doi: 10.1038/s41577-019-0266-9. Nat Rev Immunol. 2020. PMID: 31892735 Review. No abstract available.
Cited by
-
Glucose metabolism in B cell malignancies: a focus on glycolysis branching pathways.Mol Oncol. 2024 Jul;18(7):1777-1794. doi: 10.1002/1878-0261.13570. Epub 2024 Jan 3. Mol Oncol. 2024. PMID: 38115544 Free PMC article. Review.
-
Molecular Mechanisms of T Helper Cell Differentiation and Functional Specialization.Immune Netw. 2023 Feb 17;23(1):e4. doi: 10.4110/in.2023.23.e4. eCollection 2023 Feb. Immune Netw. 2023. PMID: 36911803 Free PMC article. Review.
-
Toll-Like Receptor 7 Activation Enhances CD8+ T Cell Effector Functions by Promoting Cellular Glycolysis.Front Immunol. 2019 Sep 12;10:2191. doi: 10.3389/fimmu.2019.02191. eCollection 2019. Front Immunol. 2019. PMID: 31572396 Free PMC article.
-
Clonal expansion of vaccine-elicited T cells is independent of aerobic glycolysis.Sci Immunol. 2018 Sep 7;3(27):eaas9822. doi: 10.1126/sciimmunol.aas9822. Sci Immunol. 2018. PMID: 30194241 Free PMC article.
-
Vitamin B6 Metabolism Determines T Cell Anti-Tumor Responses.Front Immunol. 2022 Feb 17;13:837669. doi: 10.3389/fimmu.2022.837669. eCollection 2022. Front Immunol. 2022. PMID: 35251031 Free PMC article.
References
-
- Blagih J, Coulombe F, Vincent Emma E, Dupuy F, Galicia-Vázquez G, Yurchenko E, Raissi Thomas C, van der Windt Gerritje JW, Viollet B, Pearce Erika L, et al. The Energy Sensor AMPK Regulates T Cell Metabolic Adaptation and Effector Responses In Vivo. Immunity. 2015;42:41–54. - PubMed
-
- Chaoul N, Fayolle C, Desrues B, Oberkampf M, Tang A, Ladant D, Leclerc C. Rapamycin Impairs Antitumor CD8+ T-cell Responses and Vaccine-Induced Tumor Eradication. Cancer Research. 2015;75:3279–3291. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
