

Reconstituted B cell receptor signaling reveals carbohydrate-dependent mode of activation
- PMID: 27796362
- PMCID: PMC5087089
- DOI: 10.1038/srep36298
Reconstituted B cell receptor signaling reveals carbohydrate-dependent mode of activation
Abstract
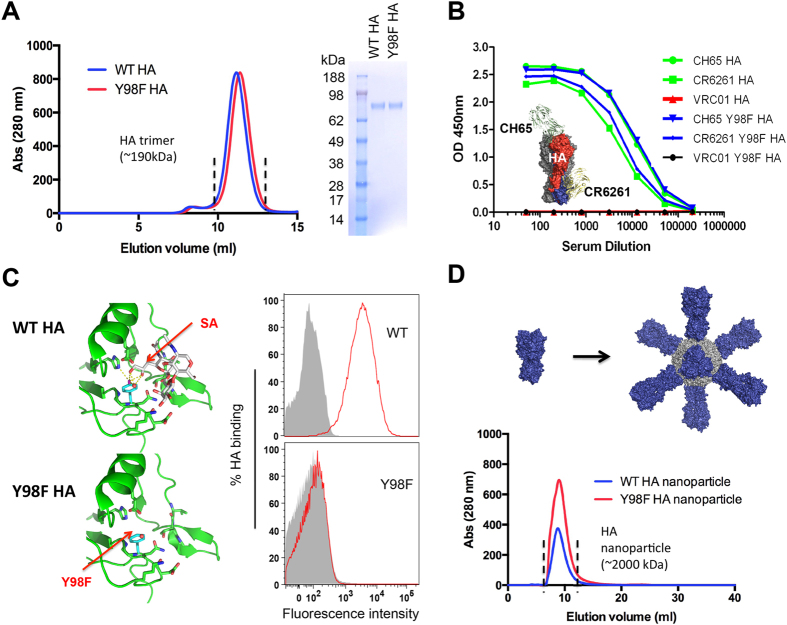
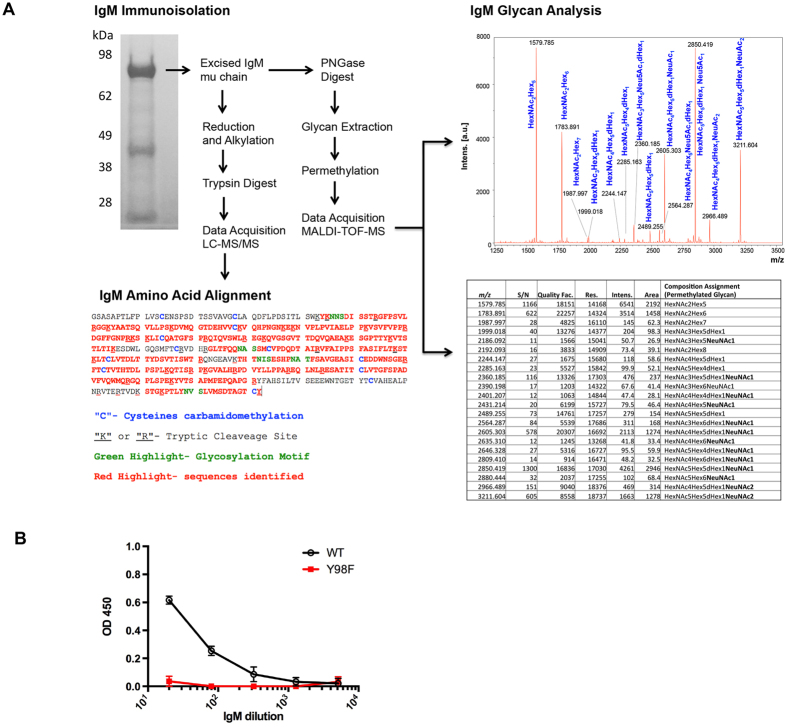
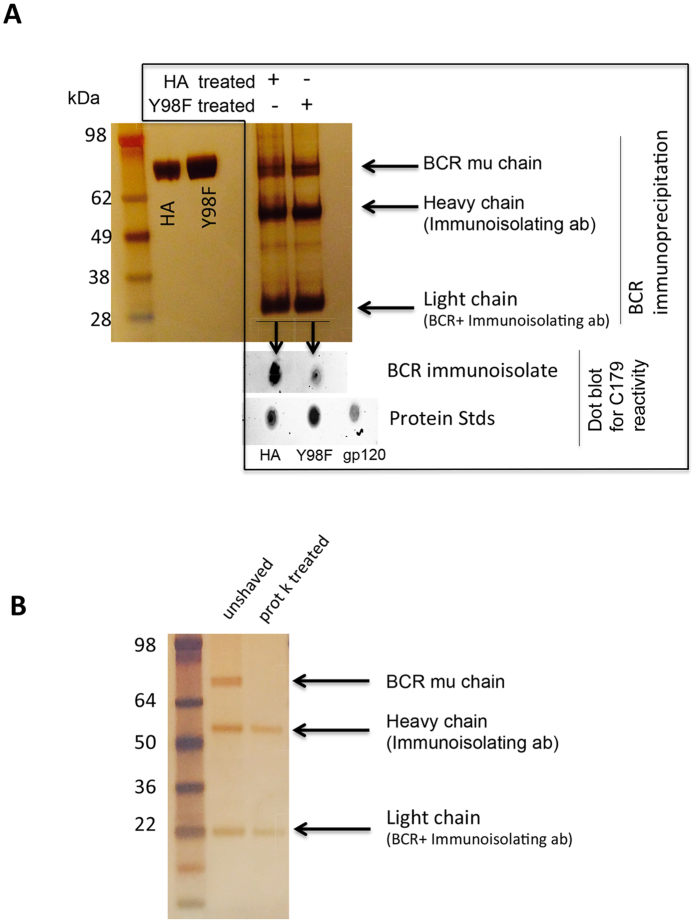
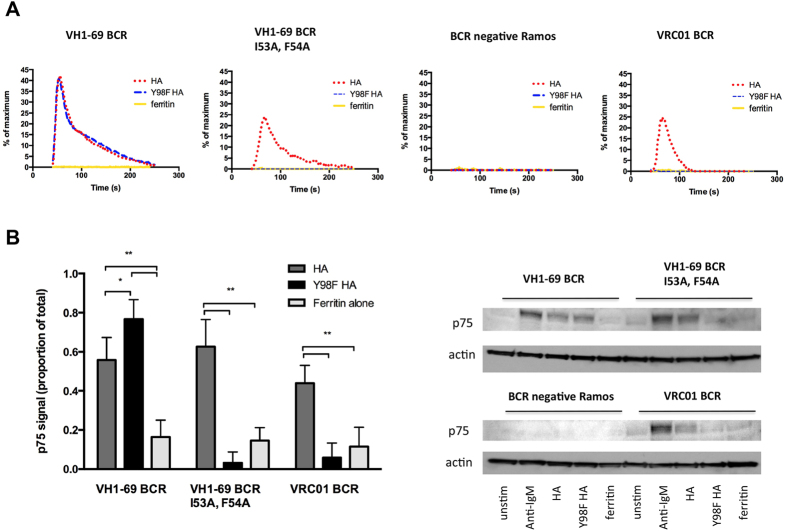
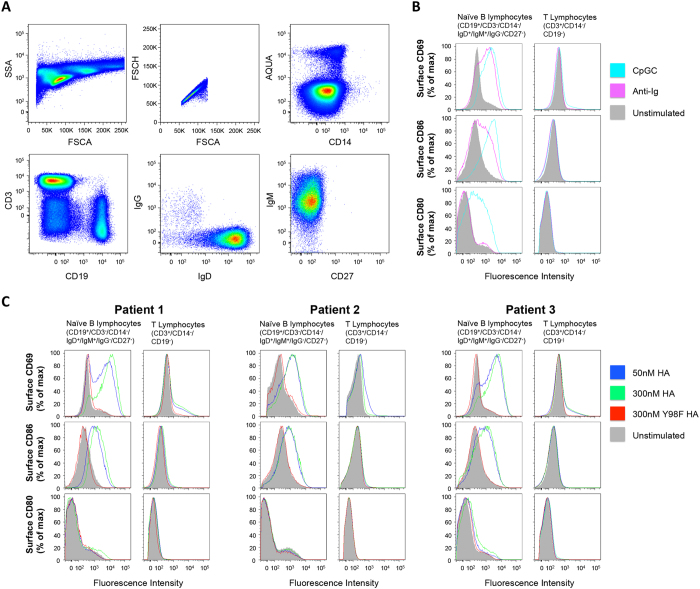
Activation of immune cells (but not B cells) with lectins is widely known. We used the structurally defined interaction between influenza hemagglutinin (HA) and its cell surface receptor sialic acid (SA) to identify a B cell receptor (BCR) activation modality that proceeded through non-cognate interactions with antigen. Using a new approach to reconstitute antigen-receptor interactions in a human reporter B cell line, we found that sequence-defined BCRs from the human germline repertoire could be triggered by both complementarity to influenza HA and a separate mode of signaling that relied on multivalent ligation of BCR sialyl-oligosaccharide. The latter suggested a new mechanism for priming naïve B cell responses and manifested as the induction of SA-dependent pan-activation by peripheral blood B cells. BCR crosslinking in the absence of complementarity is a superantigen effect induced by some microbial products to subvert production of antigen-specific immune responses. B cell superantigen activity through affinity for BCR carbohydrate is discussed.
Figures
Similar articles
-
Anti-HA Glycoforms Drive B Cell Affinity Selection and Determine Influenza Vaccine Efficacy.Cell. 2015 Jul 2;162(1):160-9. doi: 10.1016/j.cell.2015.06.026. Cell. 2015. PMID: 26140596 Free PMC article.
-
Bacterial lectin BambL acts as a B cell superantigen.Cell Mol Life Sci. 2021 Dec;78(24):8165-8186. doi: 10.1007/s00018-021-04009-z. Epub 2021 Nov 3. Cell Mol Life Sci. 2021. PMID: 34731252 Free PMC article.
-
Hemagglutinin of Influenza A Virus Antagonizes Type I Interferon (IFN) Responses by Inducing Degradation of Type I IFN Receptor 1.J Virol. 2015 Dec 16;90(5):2403-17. doi: 10.1128/JVI.02749-15. J Virol. 2015. PMID: 26676772 Free PMC article.
-
Structural and immunogenomic insights into B-cell receptor activation.J Genet Genomics. 2020 Jan 20;47(1):27-35. doi: 10.1016/j.jgg.2019.12.003. Epub 2020 Jan 9. J Genet Genomics. 2020. PMID: 32111437 Review.
-
Study B Cell Antigen Receptor Nano-Scale Organization by In Situ Fab Proximity Ligation Assay.Methods Mol Biol. 2018;1707:171-181. doi: 10.1007/978-1-4939-7474-0_12. Methods Mol Biol. 2018. PMID: 29388107 Review.
Cited by
-
Self-assembling influenza nanoparticle vaccines drive extended germinal center activity and memory B cell maturation.JCI Insight. 2020 May 21;5(10):e136653. doi: 10.1172/jci.insight.136653. JCI Insight. 2020. PMID: 32434990 Free PMC article.
-
Role of nanoscale antigen organization on B-cell activation probed using DNA origami.Nat Nanotechnol. 2020 Aug;15(8):716-723. doi: 10.1038/s41565-020-0719-0. Epub 2020 Jun 29. Nat Nanotechnol. 2020. PMID: 32601450 Free PMC article.
-
Ferritin Vaccine Platform for Animal and Zoonotic Viruses.Vaccines (Basel). 2024 Sep 27;12(10):1112. doi: 10.3390/vaccines12101112. Vaccines (Basel). 2024. PMID: 39460279 Free PMC article. Review.
-
The CH1 domain influences the expression and antigen sensing of the HIV-specific CH31 IgM-BCR and IgG-BCR.Proc Natl Acad Sci U S A. 2024 Jul 30;121(31):e2404728121. doi: 10.1073/pnas.2404728121. Epub 2024 Jul 23. Proc Natl Acad Sci U S A. 2024. PMID: 39042672 Free PMC article.
-
Engaging an HIV vaccine target through the acquisition of low B cell affinity.Nat Commun. 2023 Aug 28;14(1):5249. doi: 10.1038/s41467-023-40918-2. Nat Commun. 2023. PMID: 37640732 Free PMC article.
References
-
- Xu J. L. & Davis M. M. Diversity in the CDR3 region of V(H) is sufficient for most antibody specificities. Immunity 13, 37–45 (2000). - PubMed
-
- Davis M. M. The evolutionary and structural ‘logic’ of antigen receptor diversity. Seminars in immunology 16, 239–243 (2004). - PubMed
-
- Yang J. & Reth M. Receptor Dissociation and B-Cell Activation. Current topics in microbiology and immunology 393, 27–43 (2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
