

Dynamics of Human and Viral RNA Methylation during Zika Virus Infection
- PMID: 27773536
- PMCID: PMC5155635
- DOI: 10.1016/j.chom.2016.10.002
Dynamics of Human and Viral RNA Methylation during Zika Virus Infection
Abstract
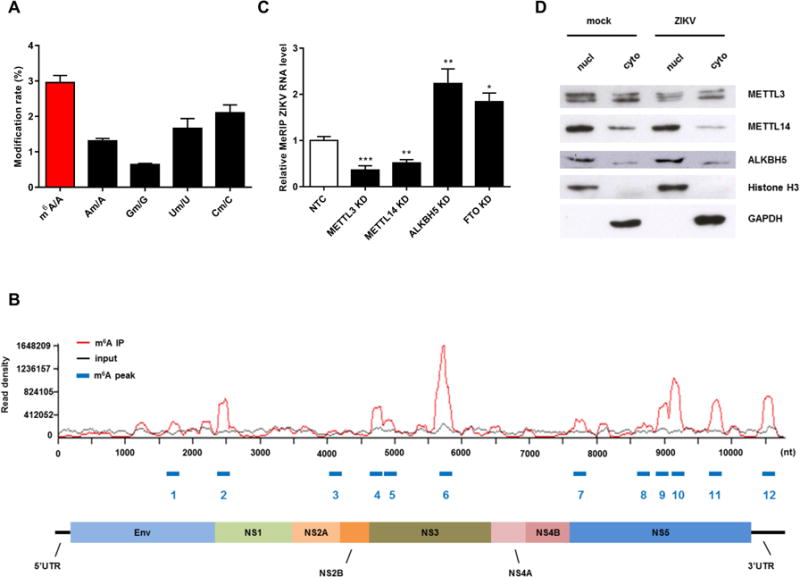
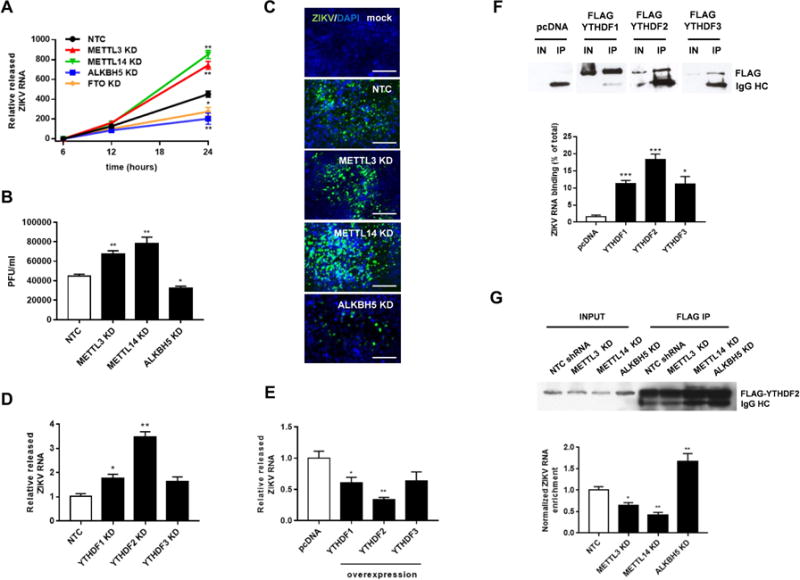
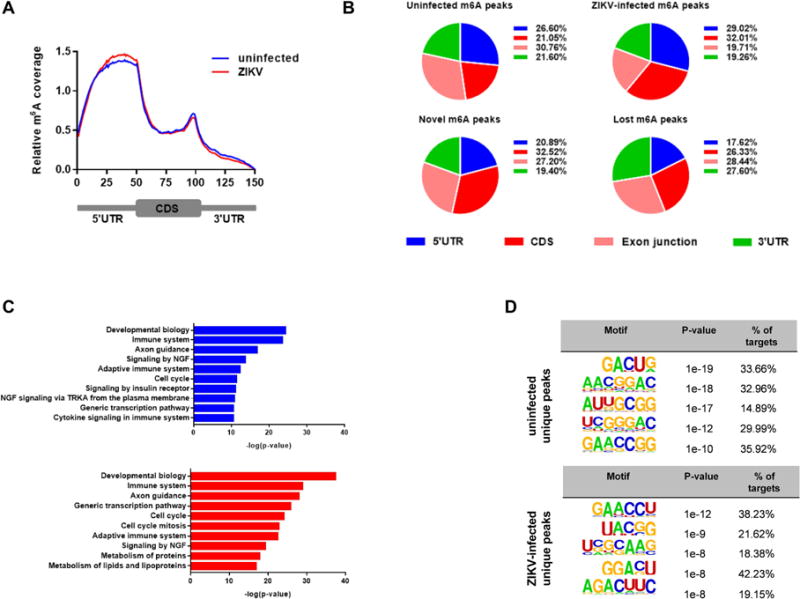
Infection with the flavivirus Zika (ZIKV) causes neurological, immunological, and developmental defects through incompletely understood mechanisms. We report that ZIKV infection affects viral and human RNAs by altering the topology and function of N6-adenosine methylation (m6A), a modification affecting RNA structure and function. m6A nucleosides are abundant in ZIKV RNA, with twelve m6A peaks identified across full-length ZIKV RNA. m6A in ZIKV RNA is controlled by host methyltransferases METTL3 and METTL14 and demethylases ALKBH5 and FTO, and knockdown of methyltransferases increases, while silencing demethylases decreases, ZIKV production. YTHDF family proteins, which regulate the stability of m6A-modified RNA, bind to ZIKV RNA, and their silencing increases ZIKV replication. Profiling of the m6A methylome of host mRNAs reveals that ZIKV infection alters m6A location in mRNAs, methylation motifs, and target genes modified by methyltransferases. Our results identify a mechanism by which ZIKV interacts with and alters host cell functions.
Keywords: 2′-O methylation; Zika virus; m(6)A methylation; methyltransferases.
Copyright © 2016 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare that they have no conflicts of interest.
Figures
Similar articles
-
METTL3 and ALKBH5 oppositely regulate m6A modification of TFEB mRNA, which dictates the fate of hypoxia/reoxygenation-treated cardiomyocytes.Autophagy. 2019 Aug;15(8):1419-1437. doi: 10.1080/15548627.2019.1586246. Epub 2019 Mar 17. Autophagy. 2019. PMID: 30870073 Free PMC article.
-
Profiling of N6-Methyladenosine in Zika Virus RNA and Host Cellular mRNA.Methods Mol Biol. 2019;1870:209-218. doi: 10.1007/978-1-4939-8808-2_16. Methods Mol Biol. 2019. PMID: 30539558
-
Functional RNA during Zika virus infection.Virus Res. 2018 Aug 2;254:41-53. doi: 10.1016/j.virusres.2017.08.015. Epub 2017 Aug 31. Virus Res. 2018. PMID: 28864425 Review.
-
m6A modification impacts hepatic drug and lipid metabolism properties by regulating carboxylesterase 2.Biochem Pharmacol. 2021 Nov;193:114766. doi: 10.1016/j.bcp.2021.114766. Epub 2021 Sep 16. Biochem Pharmacol. 2021. PMID: 34536357
-
RNA m6A modification, signals for degradation or stabilisation?Biochem Soc Trans. 2024 Apr 24;52(2):707-717. doi: 10.1042/BST20230574. Biochem Soc Trans. 2024. PMID: 38629637 Free PMC article. Review.
Cited by
-
Roles of RNA m6A modifications in plant-virus interactions.Stress Biol. 2023 Dec 18;3(1):57. doi: 10.1007/s44154-023-00133-x. Stress Biol. 2023. PMID: 38105385 Free PMC article.
-
NSUN2-mediated m5C modification of HBV RNA positively regulates HBV replication.PLoS Pathog. 2023 Dec 4;19(12):e1011808. doi: 10.1371/journal.ppat.1011808. eCollection 2023 Dec. PLoS Pathog. 2023. PMID: 38048324 Free PMC article.
-
HIV-1 Replication Benefits from the RNA Epitranscriptomic Code.J Mol Biol. 2019 Dec 6;431(24):5032-5038. doi: 10.1016/j.jmb.2019.09.021. Epub 2019 Oct 15. J Mol Biol. 2019. PMID: 31626810 Free PMC article. Review.
-
RNA m6 A modification enzymes shape innate responses to DNA by regulating interferon β.Genes Dev. 2018 Dec 1;32(23-24):1472-1484. doi: 10.1101/gad.319475.118. Epub 2018 Nov 21. Genes Dev. 2018. PMID: 30463905 Free PMC article.
-
KIAA1429/VIRMA promotes breast cancer progression by m6 A-dependent cytosolic HAS2 stabilization.EMBO Rep. 2023 Oct 9;24(10):e55506. doi: 10.15252/embr.202255506. Epub 2023 Sep 14. EMBO Rep. 2023. PMID: 37705505 Free PMC article.
References
-
- Brasil P, Pereira JP, Jr, Raja Gabaglia C, Damasceno L, Wakimoto M, Ribeiro Nogueira RM, Carvalho de Sequeira P, Machado Siqueira A, Abreu de Carvalho LM, Cotrim da Cunha D, et al. Zika Virus Infection in Pregnant Women in Rio de Janeiro – Preliminary Report. The New England journal of medicine. 2016 doi: 10.1056/NEJMoa1602412. - DOI - PMC - PubMed
-
- Broutet N, Krauer F, Riesen M, Khalakdina A, Almiron M, Aldighieri S, Espinal M, Low N, Dye C. Zika Virus as a Cause of Neurologic Disorders. The New England journal of medicine. 2016;374:1506–1509. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
