

Acetylation of VGLL4 Regulates Hippo-YAP Signaling and Postnatal Cardiac Growth
- PMID: 27720608
- PMCID: PMC5121000
- DOI: 10.1016/j.devcel.2016.09.005
Acetylation of VGLL4 Regulates Hippo-YAP Signaling and Postnatal Cardiac Growth
Abstract
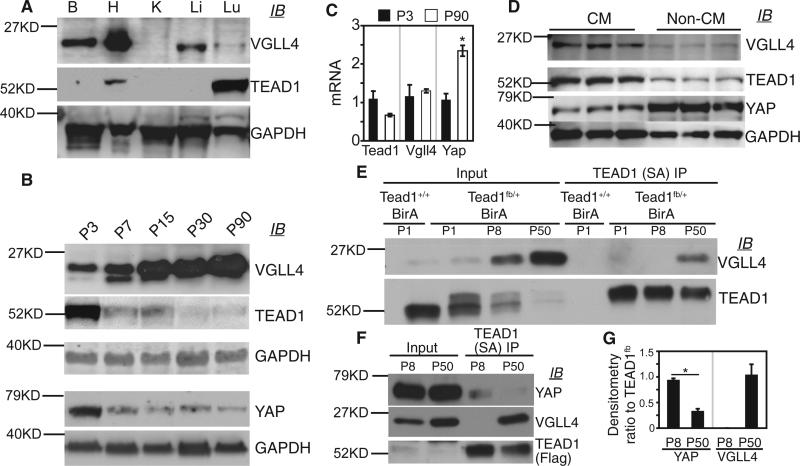
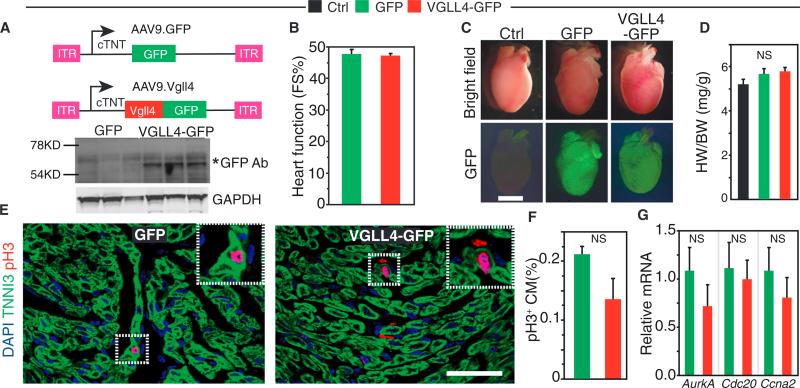
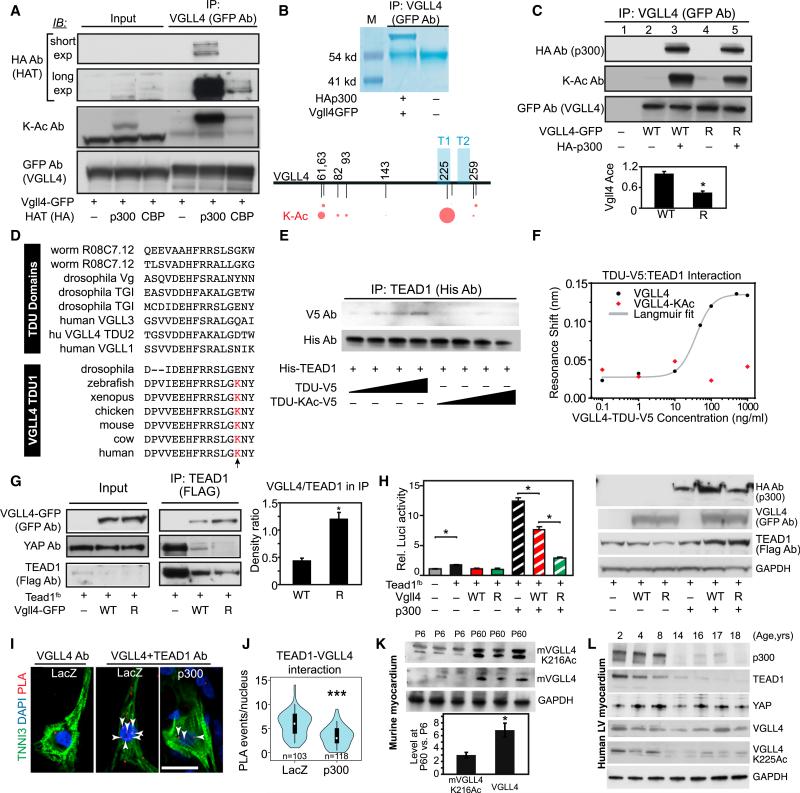
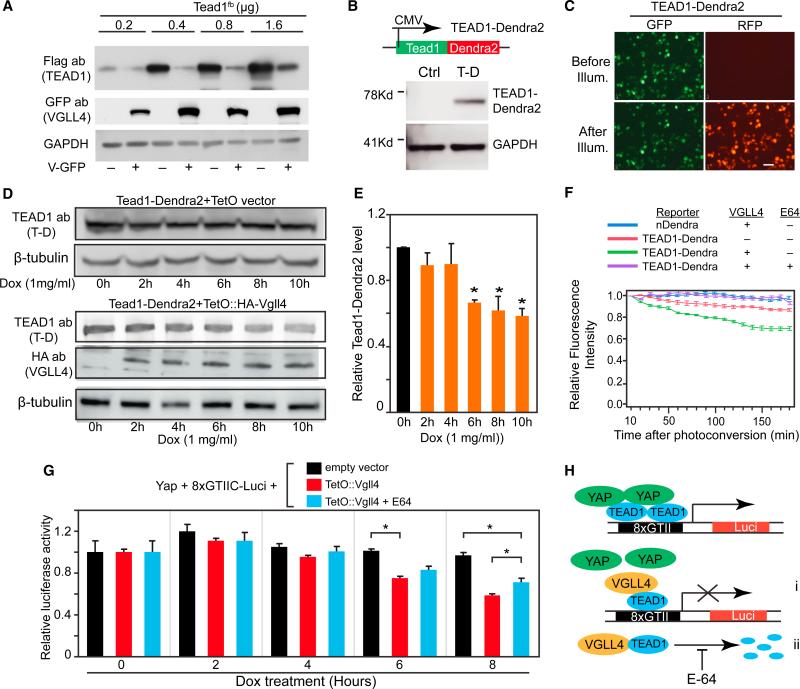
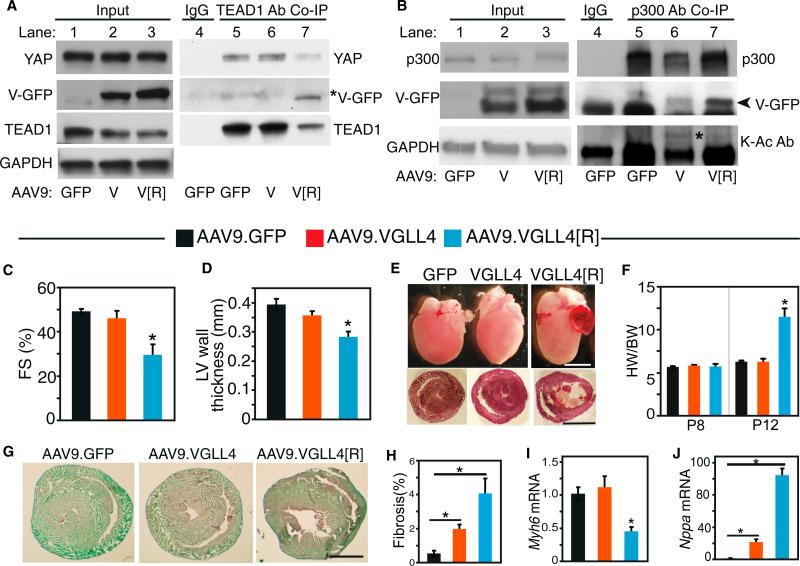
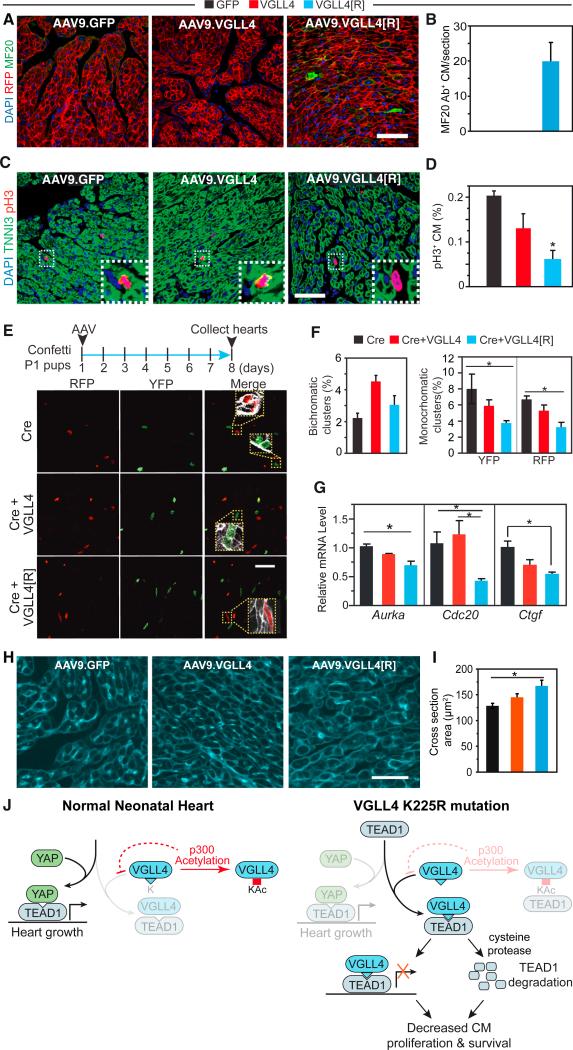
Binding of the transcriptional co-activator YAP with the transcription factor TEAD stimulates growth of the heart and other organs. YAP overexpression potently stimulates fetal cardiomyocyte (CM) proliferation, but YAP's mitogenic potency declines postnatally. While investigating factors that limit YAP's postnatal mitogenic activity, we found that the CM-enriched TEAD1 binding protein VGLL4 inhibits CM proliferation by inhibiting TEAD1-YAP interaction and by targeting TEAD1 for degradation. Importantly, VGLL4 acetylation at lysine 225 negatively regulated its binding to TEAD1. This developmentally regulated acetylation event critically governs postnatal heart growth, since overexpression of an acetylation-refractory VGLL4 mutant enhanced TEAD1 degradation, limited neonatal CM proliferation, and caused CM necrosis. Our study defines an acetylation-mediated, VGLL4-dependent switch that regulates TEAD stability and YAP-TEAD activity. These insights may improve targeted modulation of TEAD-YAP activity in applications from cardiac regeneration to cancer.
Keywords: Hippo-YAP pathway; TEAD1; VGLL4; acetylation; cardiac; cardiomyocyte; degradation; necrosis; proliferation.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Hippo Deficiency Leads to Cardiac Dysfunction Accompanied by Cardiomyocyte Dedifferentiation During Pressure Overload.Circ Res. 2019 Jan 18;124(2):292-305. doi: 10.1161/CIRCRESAHA.118.314048. Circ Res. 2019. PMID: 30582455 Free PMC article.
-
VGLL4 Protects against Oxidized-LDL-Induced Endothelial Cell Dysfunction and Inflammation by Activating Hippo-YAP/TEAD1 Signaling Pathway.Mediators Inflamm. 2020 Dec 29;2020:8292173. doi: 10.1155/2020/8292173. eCollection 2020. Mediators Inflamm. 2020. Retraction in: Mediators Inflamm. 2022 Apr 28;2022:9851919. doi: 10.1155/2022/9851919 PMID: 33456372 Free PMC article. Retracted.
-
The PDZ-binding motif of Yes-associated protein is required for its co-activation of TEAD-mediated CTGF transcription and oncogenic cell transforming activity.Biochem Biophys Res Commun. 2014 Jan 17;443(3):917-23. doi: 10.1016/j.bbrc.2013.12.100. Epub 2013 Dec 28. Biochem Biophys Res Commun. 2014. PMID: 24380865
-
The Hippo Pathway and YAP/TAZ-TEAD Protein-Protein Interaction as Targets for Regenerative Medicine and Cancer Treatment.J Med Chem. 2015 Jun 25;58(12):4857-73. doi: 10.1021/jm501615v. Epub 2015 Mar 11. J Med Chem. 2015. PMID: 25719868 Review.
-
Harnessing Hippo in the heart: Hippo/Yap signaling and applications to heart regeneration and rejuvenation.Stem Cell Res. 2014 Nov;13(3 Pt B):571-81. doi: 10.1016/j.scr.2014.04.010. Epub 2014 May 6. Stem Cell Res. 2014. PMID: 24881775 Free PMC article. Review.
Cited by
-
YAP/TAZ as mechanobiological signaling pathway in cardiovascular physiological regulation and pathogenesis.Mechanobiol Med. 2024 Dec;2(4):100085. doi: 10.1016/j.mbm.2024.100085. Epub 2024 Aug 9. Mechanobiol Med. 2024. PMID: 39281415 Free PMC article.
-
An overview of signaling pathways regulating YAP/TAZ activity.Cell Mol Life Sci. 2021 Jan;78(2):497-512. doi: 10.1007/s00018-020-03579-8. Epub 2020 Aug 3. Cell Mol Life Sci. 2021. PMID: 32748155 Free PMC article. Review.
-
Selective YAP activation in Procr cells is essential for ovarian stem/progenitor expansion and epithelium repair.Elife. 2022 Mar 14;11:e75449. doi: 10.7554/eLife.75449. Elife. 2022. PMID: 35285801 Free PMC article.
-
YAP/TEAD1 Complex Is a Default Repressor of Cardiac Toll-Like Receptor Genes.Int J Mol Sci. 2021 Jun 22;22(13):6649. doi: 10.3390/ijms22136649. Int J Mol Sci. 2021. PMID: 34206257 Free PMC article.
-
Vestigial like 4 regulates the adipogenesis of classical brown adipose tissue.bioRxiv [Preprint]. 2024 Jul 27:2024.07.09.602788. doi: 10.1101/2024.07.09.602788. bioRxiv. 2024. PMID: 39026854 Free PMC article. Preprint.
References
-
- Camargo FD, Gokhale S, Johnnidis JB, Fu D, Bell GW, Jaenisch R, Brummelkamp TR. YAP1 increases organ size and expands un-differentiated progenitor cells. Curr. Biol. 2007;17:2054–2060. - PubMed
-
- Chan HM, La Thangue NB. p300/CBP proteins: HATs for transcriptional bridges and scaffolds. J. Cell Sci. 2001;114:2363–2373. - PubMed
-
- Chen HH, Mullett SJ, Stewart AF. Vgl-4, a novel member of the vestigial-like family of transcription cofactors, regulates alpha1-adrenergic activation of gene expression in cardiac myocytes. J. Biol. Chem. 2004;279:30800–30806. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
