

Alternative polyadenylation of mRNA precursors
- PMID: 27677860
- PMCID: PMC5483950
- DOI: 10.1038/nrm.2016.116
Alternative polyadenylation of mRNA precursors
Abstract
Alternative polyadenylation (APA) is an RNA-processing mechanism that generates distinct 3' termini on mRNAs and other RNA polymerase II transcripts. It is widespread across all eukaryotic species and is recognized as a major mechanism of gene regulation. APA exhibits tissue specificity and is important for cell proliferation and differentiation. In this Review, we discuss the roles of APA in diverse cellular processes, including mRNA metabolism, protein diversification and protein localization, and more generally in gene regulation. We also discuss the molecular mechanisms underlying APA, such as variation in the concentration of core processing factors and RNA-binding proteins, as well as transcription-based regulation.
Figures
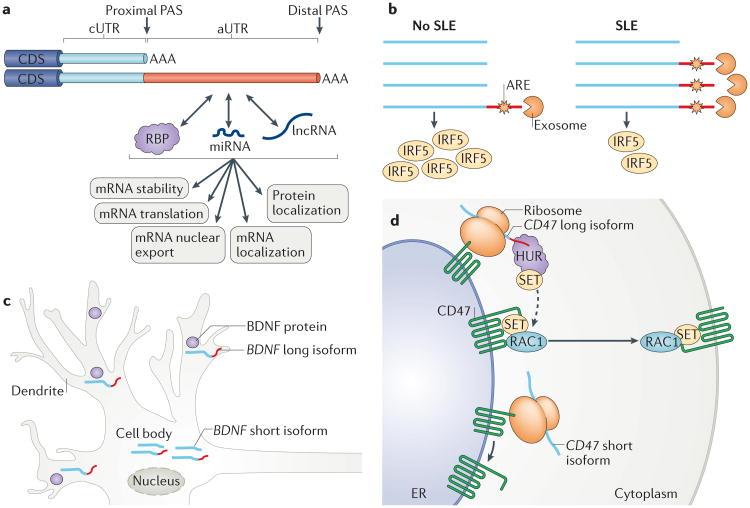
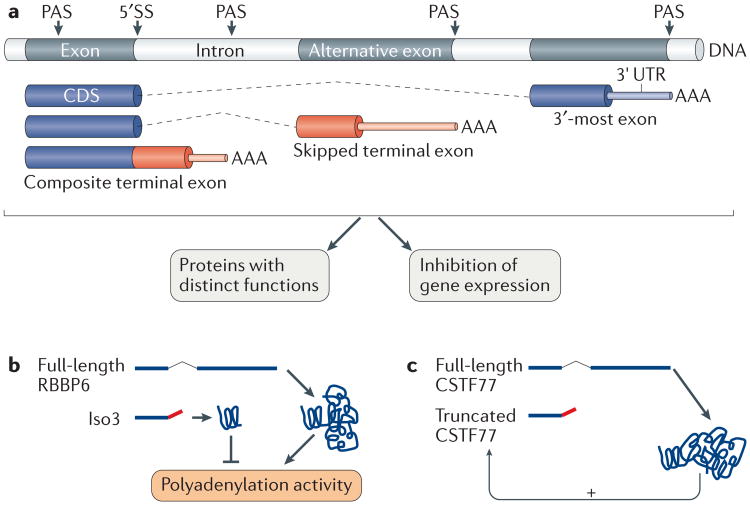
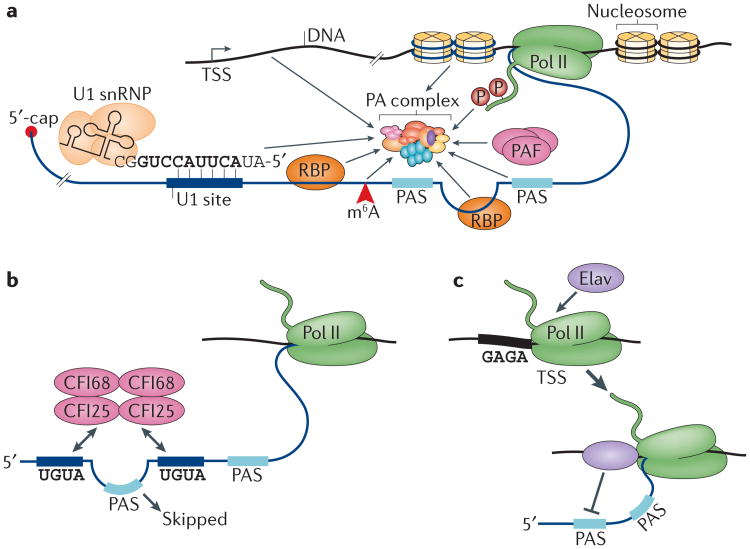
Similar articles
-
U1 snRNP telescripting: molecular mechanisms and beyond.RNA Biol. 2021 Nov;18(11):1512-1523. doi: 10.1080/15476286.2021.1872963. Epub 2021 Jan 15. RNA Biol. 2021. PMID: 33416026 Free PMC article. Review.
-
Implications of polyadenylation in health and disease.Nucleus. 2014;5(6):508-19. doi: 10.4161/nucl.36360. Epub 2014 Oct 31. Nucleus. 2014. PMID: 25484187 Free PMC article. Review.
-
Alternative polyadenylation: methods, mechanism, function, and role in cancer.J Exp Clin Cancer Res. 2021 Feb 1;40(1):51. doi: 10.1186/s13046-021-01852-7. J Exp Clin Cancer Res. 2021. PMID: 33526057 Free PMC article. Review.
-
Alternative polyadenylation and RNA-binding proteins.J Mol Endocrinol. 2016 Aug;57(2):F29-34. doi: 10.1530/JME-16-0070. Epub 2016 May 20. J Mol Endocrinol. 2016. PMID: 27208003 Review.
-
The influence of 5' and 3' end structures on pre-mRNA metabolism.J Cell Sci Suppl. 1995;19:13-9. doi: 10.1242/jcs.1995.supplement_19.2. J Cell Sci Suppl. 1995. PMID: 8655642 Review.
Cited by
-
Nuclear m6 A reader YTHDC1 suppresses proximal alternative polyadenylation sites by interfering with the 3' processing machinery.EMBO Rep. 2022 Nov 7;23(11):e54686. doi: 10.15252/embr.202254686. Epub 2022 Sep 12. EMBO Rep. 2022. PMID: 36094741 Free PMC article.
-
Diverse cell-specific patterns of alternative polyadenylation in Drosophila.Nat Commun. 2022 Sep 13;13(1):5372. doi: 10.1038/s41467-022-32305-0. Nat Commun. 2022. PMID: 36100597 Free PMC article.
-
ipaQTL-atlas: an atlas of intronic polyadenylation quantitative trait loci across human tissues.Nucleic Acids Res. 2023 Jan 6;51(D1):D1046-D1052. doi: 10.1093/nar/gkac736. Nucleic Acids Res. 2023. PMID: 36043442 Free PMC article.
-
An atlas of alternative polyadenylation quantitative trait loci contributing to complex trait and disease heritability.Nat Genet. 2021 Jul;53(7):994-1005. doi: 10.1038/s41588-021-00864-5. Epub 2021 May 13. Nat Genet. 2021. PMID: 33986536
-
A Generative Neural Network for Maximizing Fitness and Diversity of Synthetic DNA and Protein Sequences.Cell Syst. 2020 Jul 22;11(1):49-62.e16. doi: 10.1016/j.cels.2020.05.007. Epub 2020 Jun 25. Cell Syst. 2020. PMID: 32711843 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
