

Thermal Unfolding of the Pertussis Toxin S1 Subunit Facilitates Toxin Translocation to the Cytosol by the Mechanism of Endoplasmic Reticulum-Associated Degradation
- PMID: 27647866
- PMCID: PMC5116717
- DOI: 10.1128/IAI.00732-16
Thermal Unfolding of the Pertussis Toxin S1 Subunit Facilitates Toxin Translocation to the Cytosol by the Mechanism of Endoplasmic Reticulum-Associated Degradation
Abstract
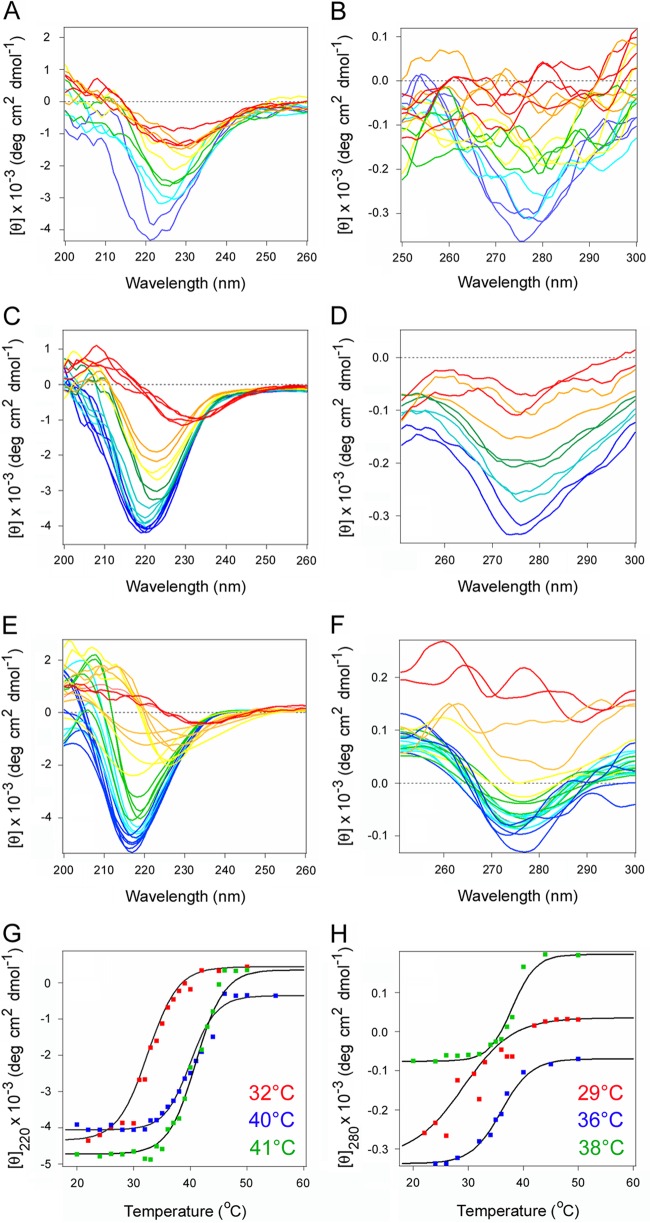
Pertussis toxin (PT) moves from the host cell surface to the endoplasmic reticulum (ER) by retrograde vesicular transport. The catalytic PTS1 subunit dissociates from the rest of the toxin in the ER and then shifts to a disordered conformation which may trigger its export to the cytosol through the quality control mechanism of ER-associated degradation (ERAD). Functional roles for toxin instability and ERAD in PTS1 translocation have not been established. We addressed these issues with the use of a surface plasmon resonance system to quantify the cytosolic pool of PTS1 from intoxicated cells. Only 3% of surface-associated PTS1 reached the host cytosol after 3 h of toxin exposure. This represented, on average, 38,000 molecules of cytosolic PTS1 per cell. Cells treated with a proteasome inhibitor contained larger quantities of cytosolic PTS1. Stabilization of the dissociated PTS1 subunit with chemical chaperones inhibited toxin export to the cytosol and blocked PT intoxication. ERAD-defective cell lines likewise exhibited reduced quantities of cytosolic PTS1 and PT resistance. These observations identify the unfolding of dissociated PTS1 as a trigger for its ERAD-mediated translocation to the cytosol.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Figures
Similar articles
-
Intracellular Trafficking and Translocation of Pertussis Toxin.Toxins (Basel). 2019 Jul 25;11(8):437. doi: 10.3390/toxins11080437. Toxins (Basel). 2019. PMID: 31349590 Free PMC article. Review.
-
The pertussis toxin S1 subunit is a thermally unstable protein susceptible to degradation by the 20S proteasome.Biochemistry. 2006 Nov 21;45(46):13734-40. doi: 10.1021/bi061175+. Biochemistry. 2006. PMID: 17105192 Free PMC article.
-
Evading the proteasome: absence of lysine residues contributes to pertussis toxin activity by evasion of proteasome degradation.Infect Immun. 2007 Jun;75(6):2946-53. doi: 10.1128/IAI.02011-06. Epub 2007 Apr 9. Infect Immun. 2007. PMID: 17420233 Free PMC article.
-
The cholera toxin A1(3) subdomain is essential for interaction with ADP-ribosylation factor 6 and full toxic activity but is not required for translocation from the endoplasmic reticulum to the cytosol.Infect Immun. 2006 Apr;74(4):2259-67. doi: 10.1128/IAI.74.4.2259-2267.2006. Infect Immun. 2006. PMID: 16552056 Free PMC article.
-
A bacterial toxin and a nonenveloped virus hijack ER-to-cytosol membrane translocation pathways to cause disease.Crit Rev Biochem Mol Biol. 2015;50(6):477-88. doi: 10.3109/10409238.2015.1085826. Epub 2015 Sep 11. Crit Rev Biochem Mol Biol. 2015. PMID: 26362261 Free PMC article. Review.
Cited by
-
Aluminum hydroxide adjuvant diverts the uptake and trafficking of genetically detoxified pertussis toxin to lysosomes in macrophages.Mol Microbiol. 2022 May;117(5):1173-1195. doi: 10.1111/mmi.14900. Epub 2022 Apr 7. Mol Microbiol. 2022. PMID: 35344242 Free PMC article.
-
Domperidone Inhibits Clostridium botulinum C2 Toxin and Bordetella pertussis Toxin.Toxins (Basel). 2023 Jun 25;15(7):412. doi: 10.3390/toxins15070412. Toxins (Basel). 2023. PMID: 37505681 Free PMC article.
-
Pharmacological targeting of host chaperones protects from pertussis toxin in vitro and in vivo.Sci Rep. 2021 Mar 8;11(1):5429. doi: 10.1038/s41598-021-84817-2. Sci Rep. 2021. PMID: 33686161 Free PMC article.
-
Toxins Utilize the Endoplasmic Reticulum-Associated Protein Degradation Pathway in Their Intoxication Process.Int J Mol Sci. 2019 Mar 15;20(6):1307. doi: 10.3390/ijms20061307. Int J Mol Sci. 2019. PMID: 30875878 Free PMC article. Review.
-
Human Peptides α-Defensin-1 and -5 Inhibit Pertussis Toxin.Toxins (Basel). 2021 Jul 11;13(7):480. doi: 10.3390/toxins13070480. Toxins (Basel). 2021. PMID: 34357952 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
