

Internetwork competition for monomers governs actin cytoskeleton organization
- PMID: 27625321
- PMCID: PMC5125073
- DOI: 10.1038/nrm.2016.106
Internetwork competition for monomers governs actin cytoskeleton organization
Abstract
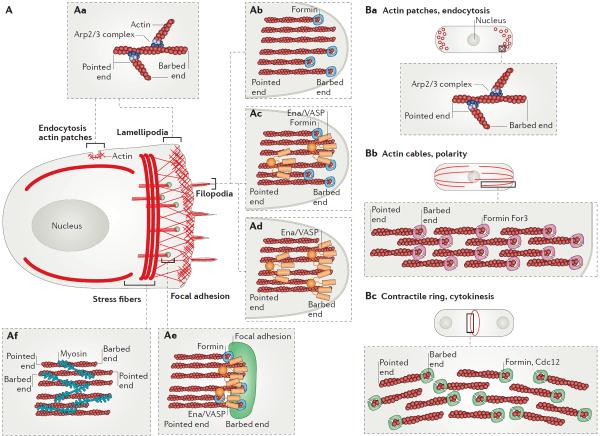
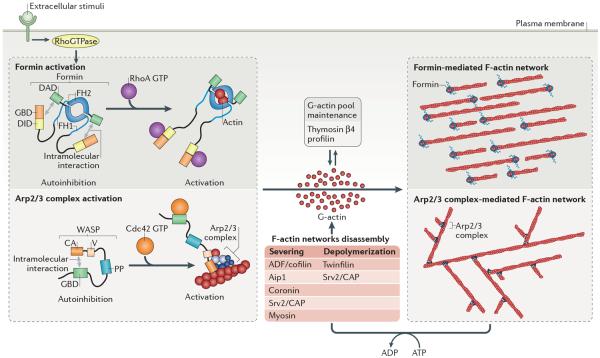
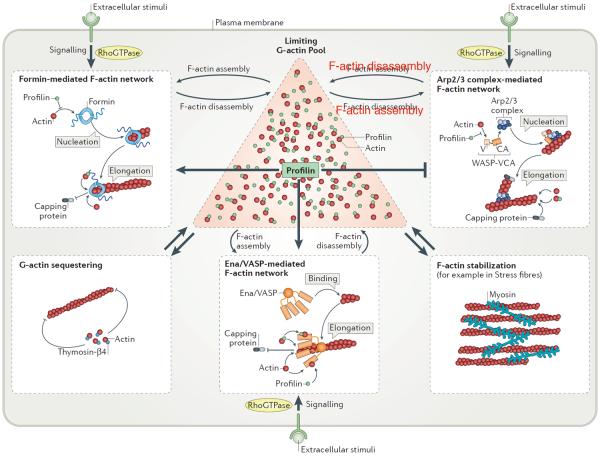
Cells precisely control the formation of dynamic actin cytoskeleton networks to coordinate fundamental processes, including motility, division, endocytosis and polarization. To support these functions, actin filament networks must be assembled, maintained and disassembled at the correct time and place, and with proper filament organization and dynamics. Regulation of the extent of filament network assembly and of filament network organization has been largely attributed to the coordinated activation of actin assembly factors through signalling cascades. Here, we discuss an intriguing model in which actin monomer availability is limiting and competition between homeostatic actin cytoskeletal networks for actin monomers is an additional crucial regulatory mechanism that influences the density and size of different actin networks, thereby contributing to the organization of the cellular actin cytoskeleton.
Figures

Similar articles
-
Actin assembly mechanisms at a glance.J Cell Sci. 2017 Oct 15;130(20):3427-3435. doi: 10.1242/jcs.206433. J Cell Sci. 2017. PMID: 29032357 Review.
-
F-Actin Cytoskeleton Network Self-Organization Through Competition and Cooperation.Annu Rev Cell Dev Biol. 2020 Oct 6;36:35-60. doi: 10.1146/annurev-cellbio-032320-094706. Annu Rev Cell Dev Biol. 2020. PMID: 33021819 Free PMC article. Review.
-
Profilin regulates F-actin network homeostasis by favoring formin over Arp2/3 complex.Dev Cell. 2015 Jan 12;32(1):43-53. doi: 10.1016/j.devcel.2014.10.027. Epub 2014 Dec 24. Dev Cell. 2015. PMID: 25543282 Free PMC article.
-
Profilin Interaction with Actin Filament Barbed End Controls Dynamic Instability, Capping, Branching, and Motility.Dev Cell. 2016 Jan 25;36(2):201-14. doi: 10.1016/j.devcel.2015.12.024. Dev Cell. 2016. PMID: 26812019 Free PMC article.
-
Directed actin assembly and motility.Methods Enzymol. 2014;540:283-300. doi: 10.1016/B978-0-12-397924-7.00016-9. Methods Enzymol. 2014. PMID: 24630113
Cited by
-
Pleiotropic regulatory locus 1 maintains actin cytoskeleton integrity and cellular homeostasis to enable Arabidopsis root growth.iScience. 2024 Jun 28;27(8):110414. doi: 10.1016/j.isci.2024.110414. eCollection 2024 Aug 16. iScience. 2024. PMID: 39108734 Free PMC article.
-
A heterologous in-cell assay for investigating intermicrovillar adhesion complex interactions reveals a novel protrusion length-matching mechanism.J Biol Chem. 2020 Nov 27;295(48):16191-16206. doi: 10.1074/jbc.RA120.015929. Epub 2020 Oct 13. J Biol Chem. 2020. PMID: 33051206 Free PMC article.
-
Growth-induced collective bending and kinetic trapping of cytoskeletal filaments.bioRxiv [Preprint]. 2024 Jan 10:2024.01.09.574885. doi: 10.1101/2024.01.09.574885. bioRxiv. 2024. Update in: Cytoskeleton (Hoboken). 2024 Aug;81(8):409-419. doi: 10.1002/cm.21877 PMID: 38260433 Free PMC article. Updated. Preprint.
-
Actin stabilizing compounds show specific biological effects due to their binding mode.Sci Rep. 2019 Jul 5;9(1):9731. doi: 10.1038/s41598-019-46282-w. Sci Rep. 2019. PMID: 31278311 Free PMC article.
-
Simultaneous quantification of actin monomer and filament dynamics with modeling-assisted analysis of photoactivation.J Cell Sci. 2016 Dec 15;129(24):4633-4643. doi: 10.1242/jcs.194670. Epub 2016 Nov 9. J Cell Sci. 2016. PMID: 27831495 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
