

Allosteric Activation of SAMHD1 Protein by Deoxynucleotide Triphosphate (dNTP)-dependent Tetramerization Requires dNTP Concentrations That Are Similar to dNTP Concentrations Observed in Cycling T Cells
- PMID: 27566548
- PMCID: PMC5076810
- DOI: 10.1074/jbc.C116.751446
Allosteric Activation of SAMHD1 Protein by Deoxynucleotide Triphosphate (dNTP)-dependent Tetramerization Requires dNTP Concentrations That Are Similar to dNTP Concentrations Observed in Cycling T Cells
Abstract
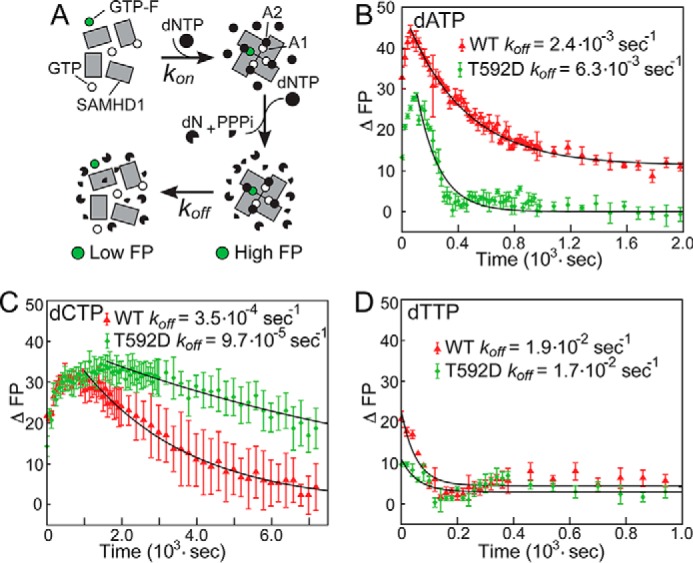
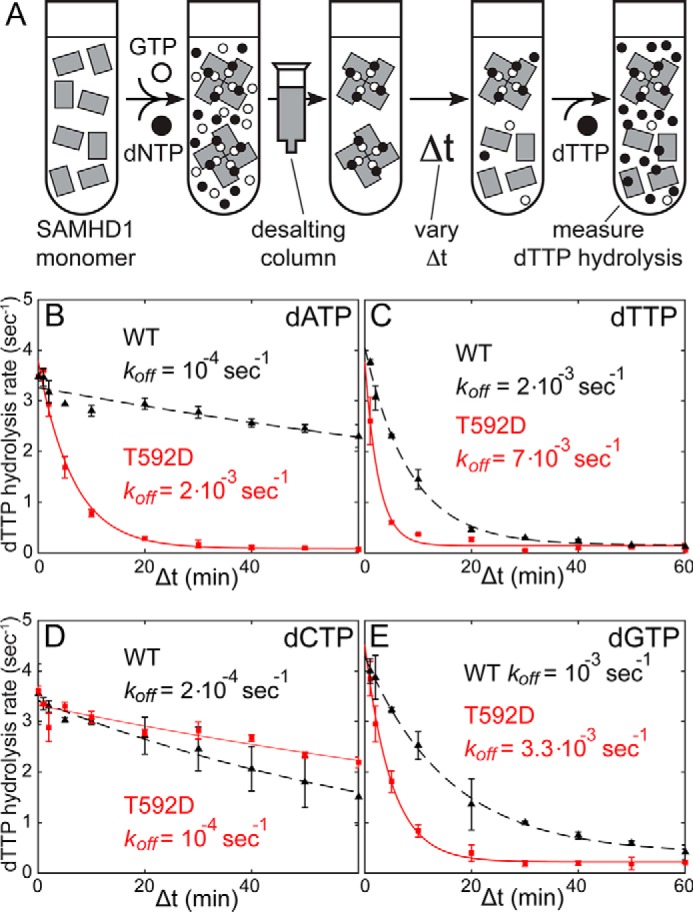
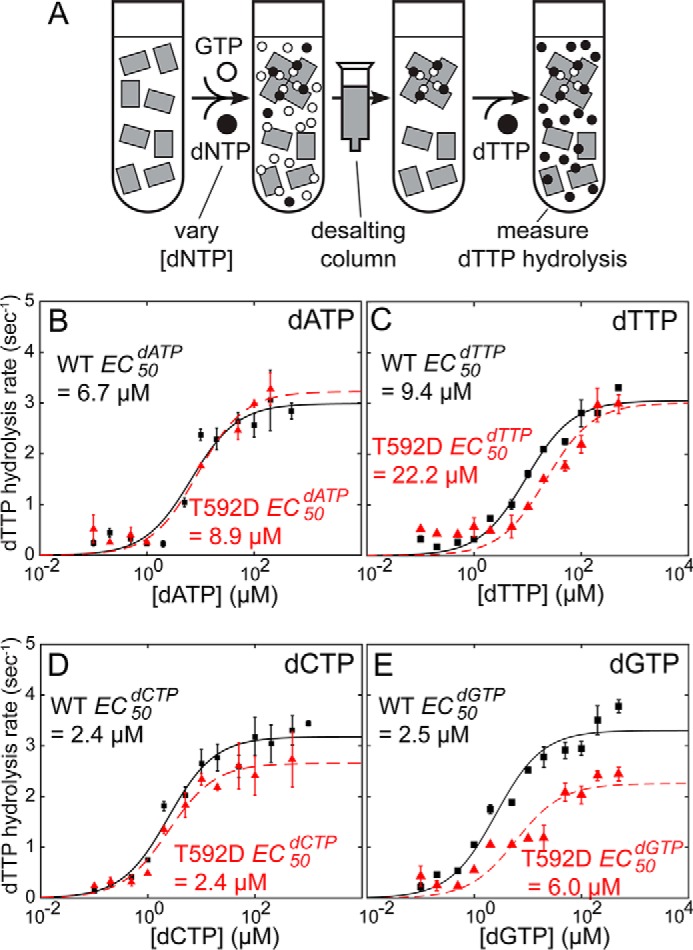
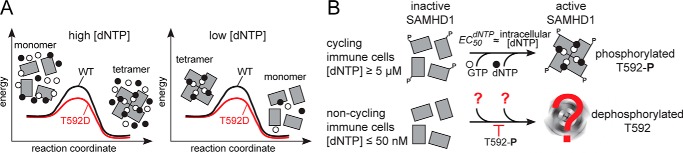
SAMHD1 is a dNTP hydrolase, whose activity is required for maintaining low dNTP concentrations in non-cycling T cells, dendritic cells, and macrophages. SAMHD1-dependent dNTP depletion is thought to impair retroviral replication in these cells, but the relationship between the dNTPase activity and retroviral restriction is not fully understood. In this study, we investigate allosteric activation of SAMHD1 by deoxynucleotide-dependent tetramerization and measure how the lifetime of the enzymatically active tetramer is affected by different dNTP ligands bound in the allosteric site. The EC50dNTP values for SAMHD1 activation by dNTPs are in the 2-20 μm range, and the half-life of the assembled tetramer after deoxynucleotide depletion varies from minutes to hours depending on what dNTP is bound in the A2 allosteric site. Comparison of the wild-type SAMHD1 and the T592D mutant reveals that the phosphomimetic mutation affects the rates of tetramer dissociation, but has no effect on the equilibrium of allosteric activation by deoxynucleotides. Collectively, our data suggest that deoxynucleotide-dependent tetramerization contributes to regulation of deoxynucleotide levels in cycling cells, whereas in non-cycling cells restrictive to retroviral replication, SAMHD1 activation is likely to be achieved through a distinct mechanism.
Keywords: autoimmune disease; human immunodeficiency virus (HIV); innate immunity; nucleoside/nucleotide metabolism; retrovirus.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
Similar articles
-
SAMHD1 Functions and Human Diseases.Viruses. 2020 Mar 31;12(4):382. doi: 10.3390/v12040382. Viruses. 2020. PMID: 32244340 Free PMC article. Review.
-
Structural basis of cellular dNTP regulation by SAMHD1.Proc Natl Acad Sci U S A. 2014 Oct 14;111(41):E4305-14. doi: 10.1073/pnas.1412289111. Epub 2014 Sep 29. Proc Natl Acad Sci U S A. 2014. PMID: 25267621 Free PMC article.
-
GTP activator and dNTP substrates of HIV-1 restriction factor SAMHD1 generate a long-lived activated state.Proc Natl Acad Sci U S A. 2014 May 6;111(18):E1843-51. doi: 10.1073/pnas.1401706111. Epub 2014 Apr 21. Proc Natl Acad Sci U S A. 2014. PMID: 24753578 Free PMC article.
-
Structural basis of allosteric activation of sterile α motif and histidine-aspartate domain-containing protein 1 (SAMHD1) by nucleoside triphosphates.J Biol Chem. 2014 Nov 21;289(47):32617-27. doi: 10.1074/jbc.M114.591958. Epub 2014 Oct 6. J Biol Chem. 2014. PMID: 25288794 Free PMC article.
-
The missing link: allostery and catalysis in the anti-viral protein SAMHD1.Biochem Soc Trans. 2019 Aug 30;47(4):1013-1027. doi: 10.1042/BST20180348. Epub 2019 Jul 11. Biochem Soc Trans. 2019. PMID: 31296733 Free PMC article. Review.
Cited by
-
SAMHD1 Functions and Human Diseases.Viruses. 2020 Mar 31;12(4):382. doi: 10.3390/v12040382. Viruses. 2020. PMID: 32244340 Free PMC article. Review.
-
SAMHD1 phosphorylation and cytoplasmic relocalization after human cytomegalovirus infection limits its antiviral activity.PLoS Pathog. 2020 Sep 28;16(9):e1008855. doi: 10.1371/journal.ppat.1008855. eCollection 2020 Sep. PLoS Pathog. 2020. PMID: 32986788 Free PMC article.
-
The ability of SAMHD1 to block HIV-1 but not SIV requires expression of MxB.Virology. 2019 May;531:260-268. doi: 10.1016/j.virol.2019.03.018. Epub 2019 Mar 30. Virology. 2019. PMID: 30959264 Free PMC article.
-
SAMHD1: Recurring roles in cell cycle, viral restriction, cancer, and innate immunity.Autoimmunity. 2018 May;51(3):96-110. doi: 10.1080/08916934.2018.1454912. Epub 2018 Mar 27. Autoimmunity. 2018. PMID: 29583030 Free PMC article. Review.
-
Functionality of Redox-Active Cysteines Is Required for Restriction of Retroviral Replication by SAMHD1.Cell Rep. 2018 Jul 24;24(4):815-823. doi: 10.1016/j.celrep.2018.06.090. Cell Rep. 2018. PMID: 30044979 Free PMC article.
References
-
- Berger A., Sommer A. F., Zwarg J., Hamdorf M., Welzel K., Esly N., Panitz S., Reuter A., Ramos I., Jatiani A., Mulder L. C., Fernandez-Sesma A., Rutsch F., Simon V., König R., and Flory E. (2011) SAMHD1-deficient CD14+ cells from individuals with Aicardi-Goutieres syndrome are highly susceptible to HIV-1 infection. PLoS Pathog. 7, e1002425. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
