

Rab5 and its effector FHF contribute to neuronal polarity through dynein-dependent retrieval of somatodendritic proteins from the axon
- PMID: 27559088
- PMCID: PMC5018783
- DOI: 10.1073/pnas.1601844113
Rab5 and its effector FHF contribute to neuronal polarity through dynein-dependent retrieval of somatodendritic proteins from the axon
Abstract
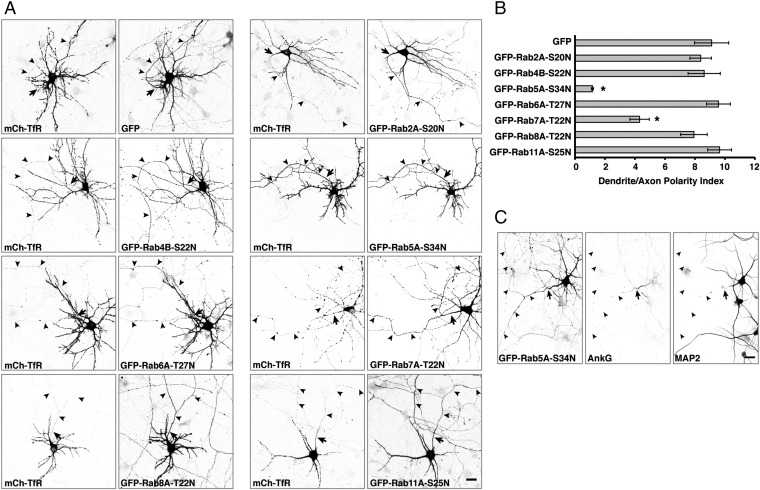
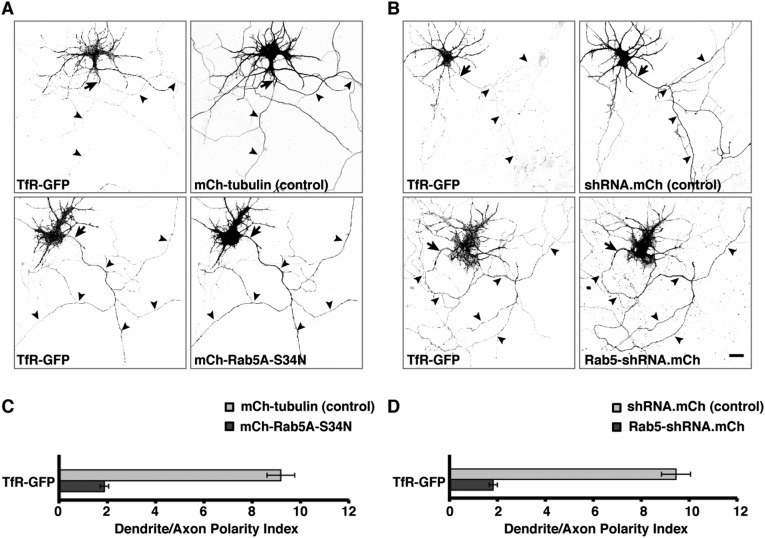
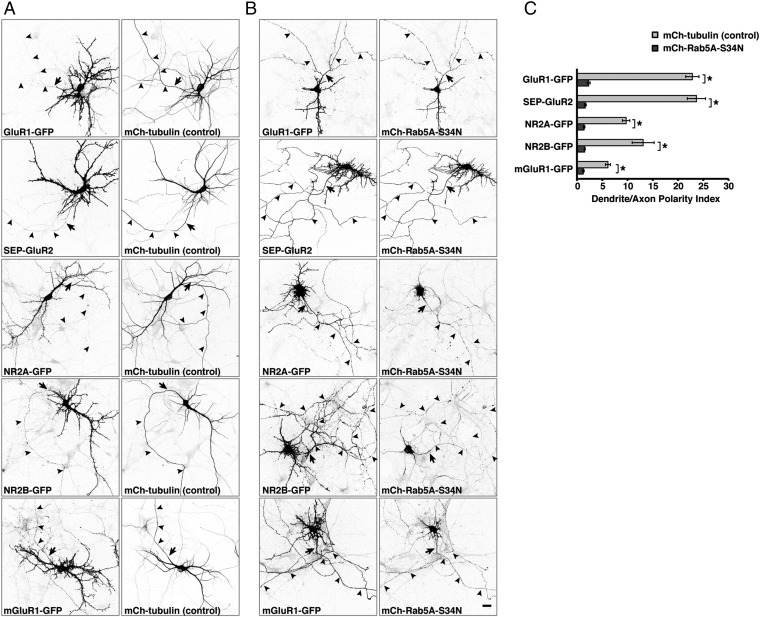
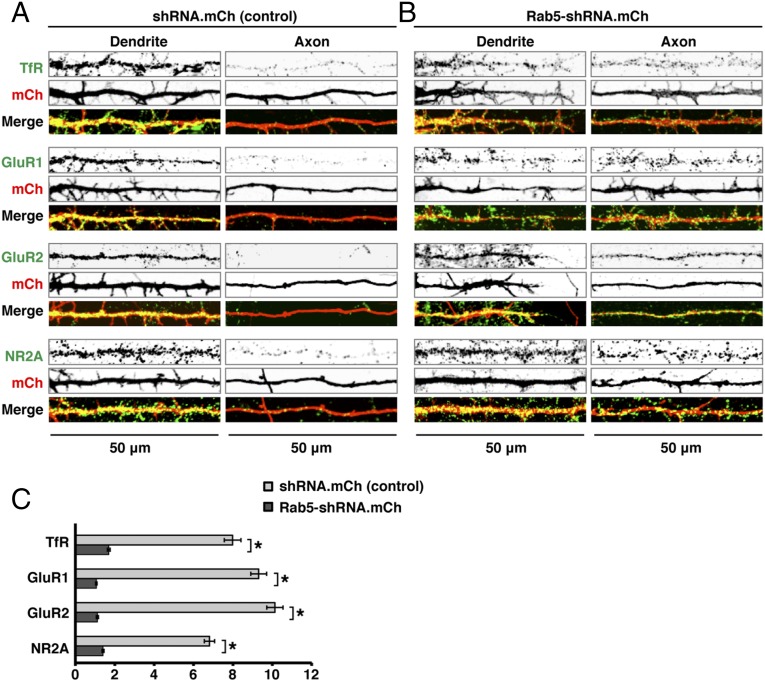
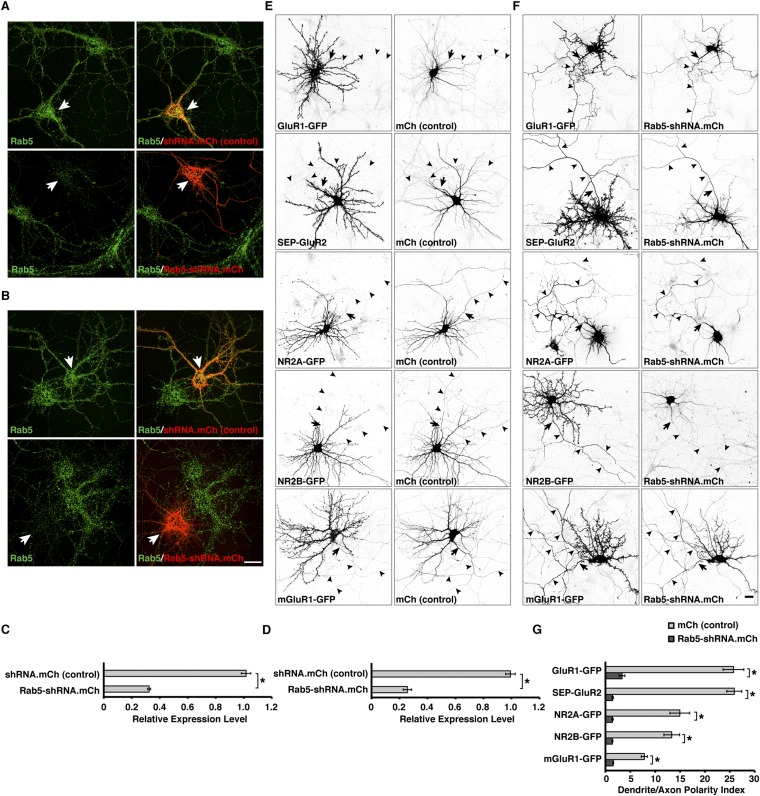
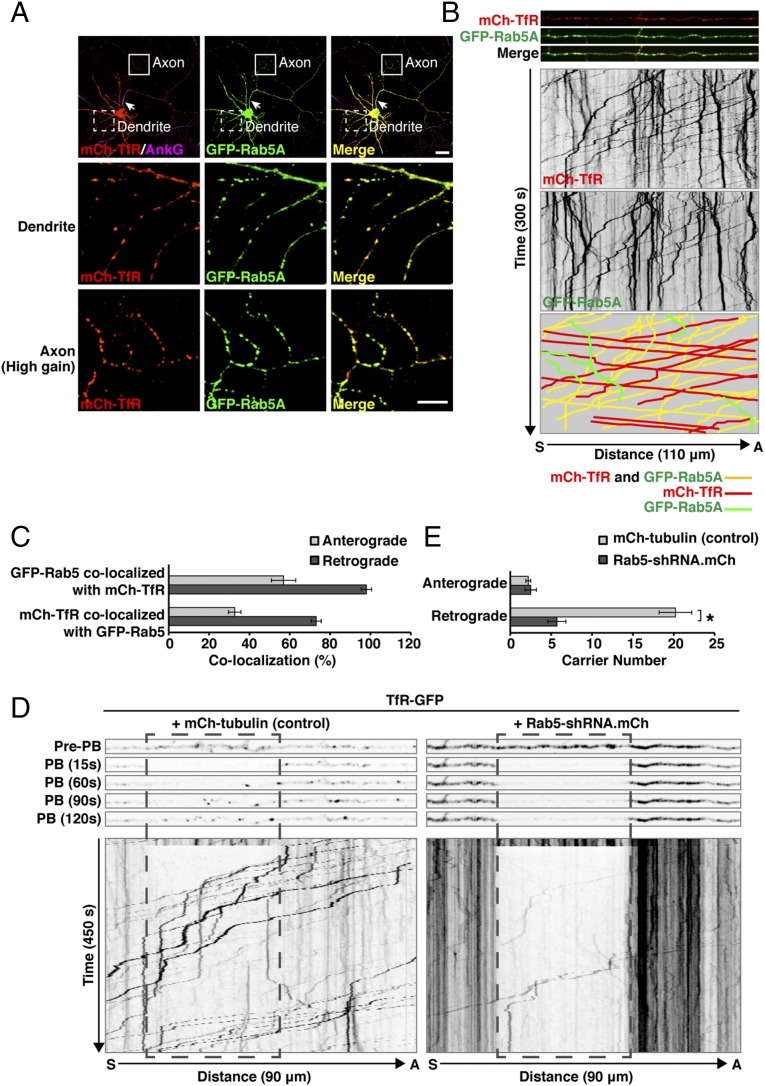
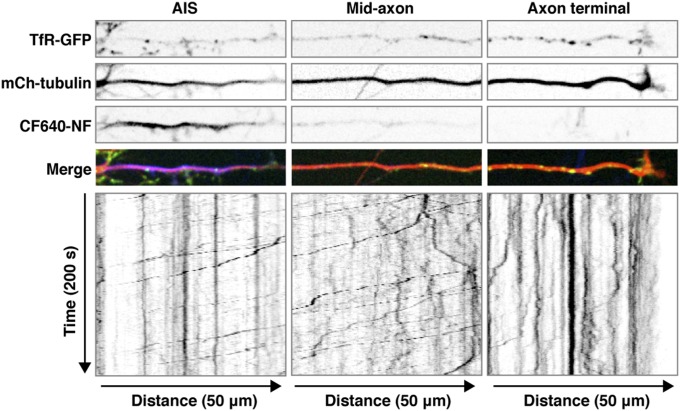
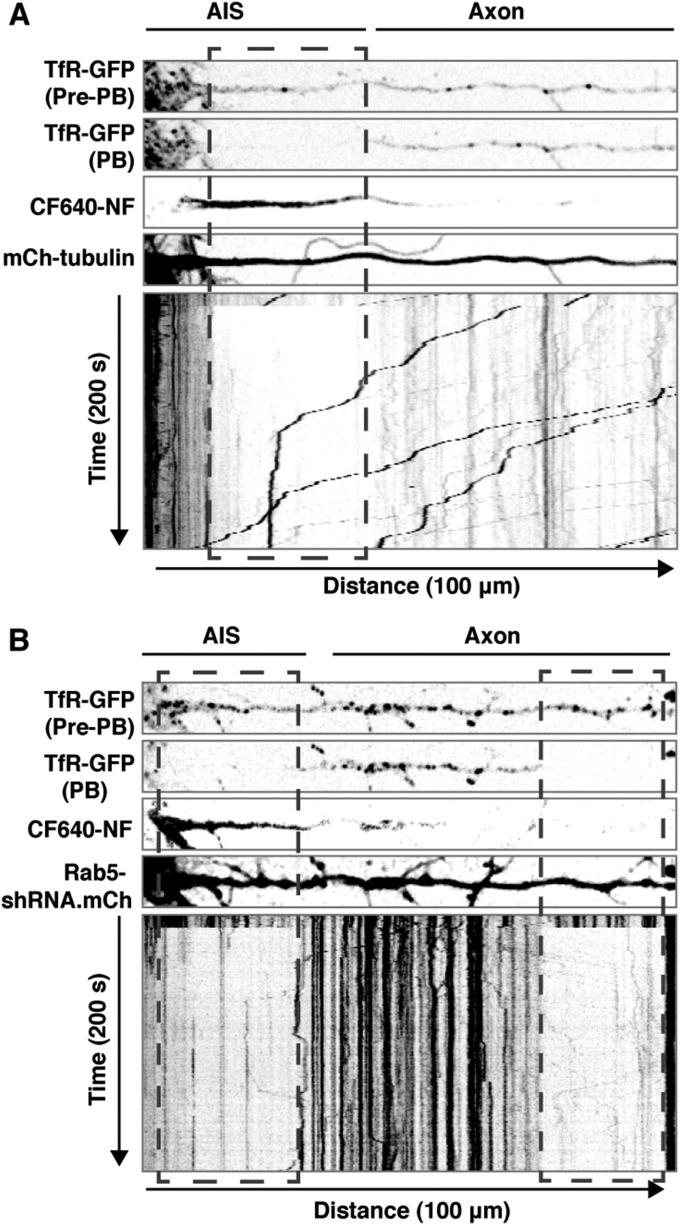
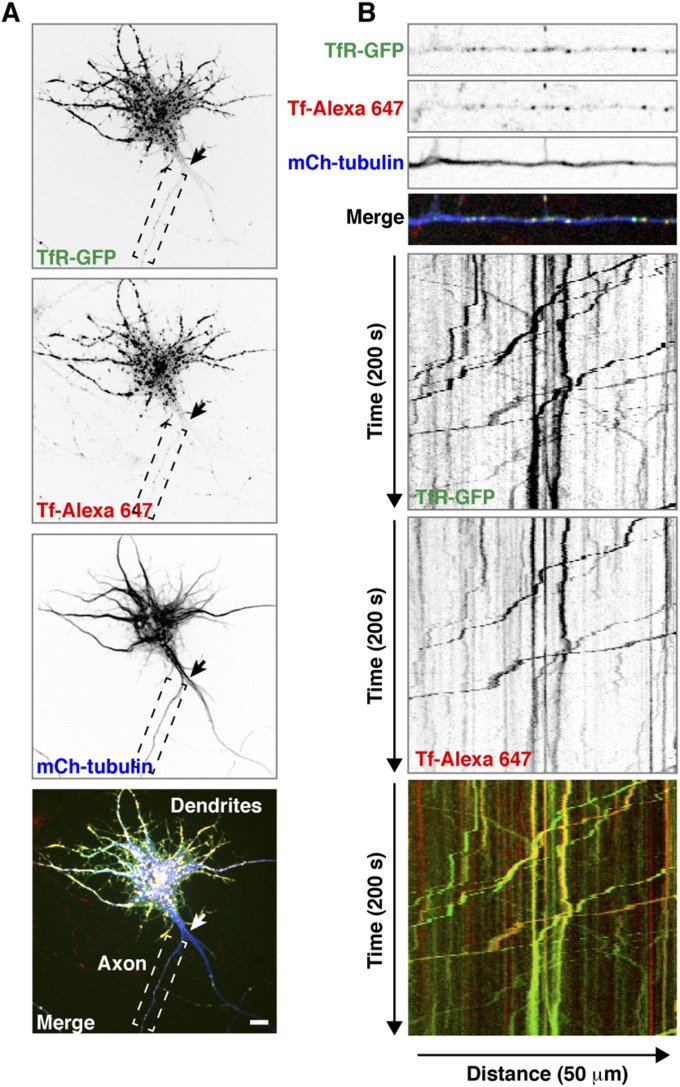
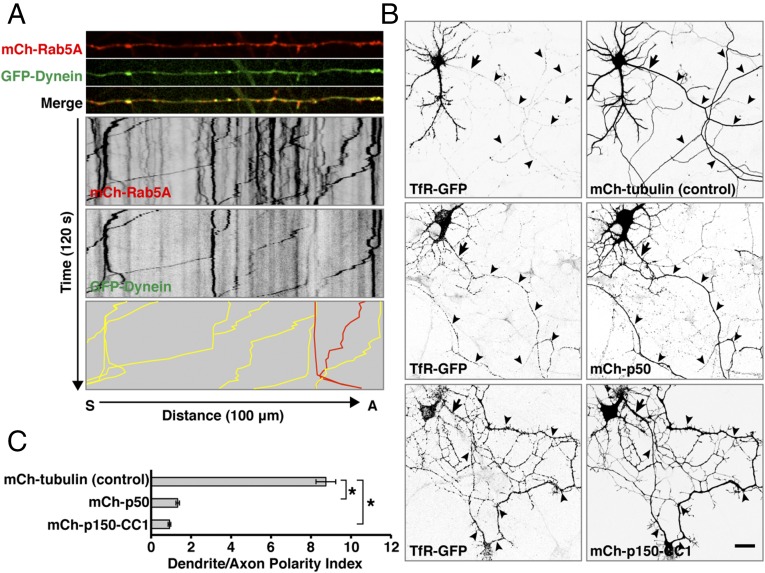
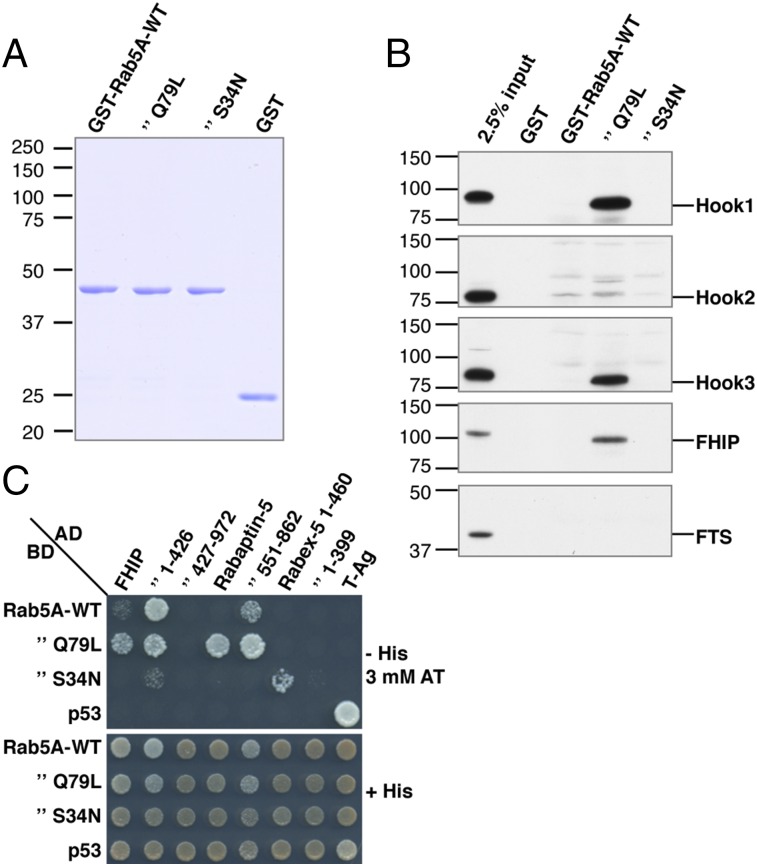
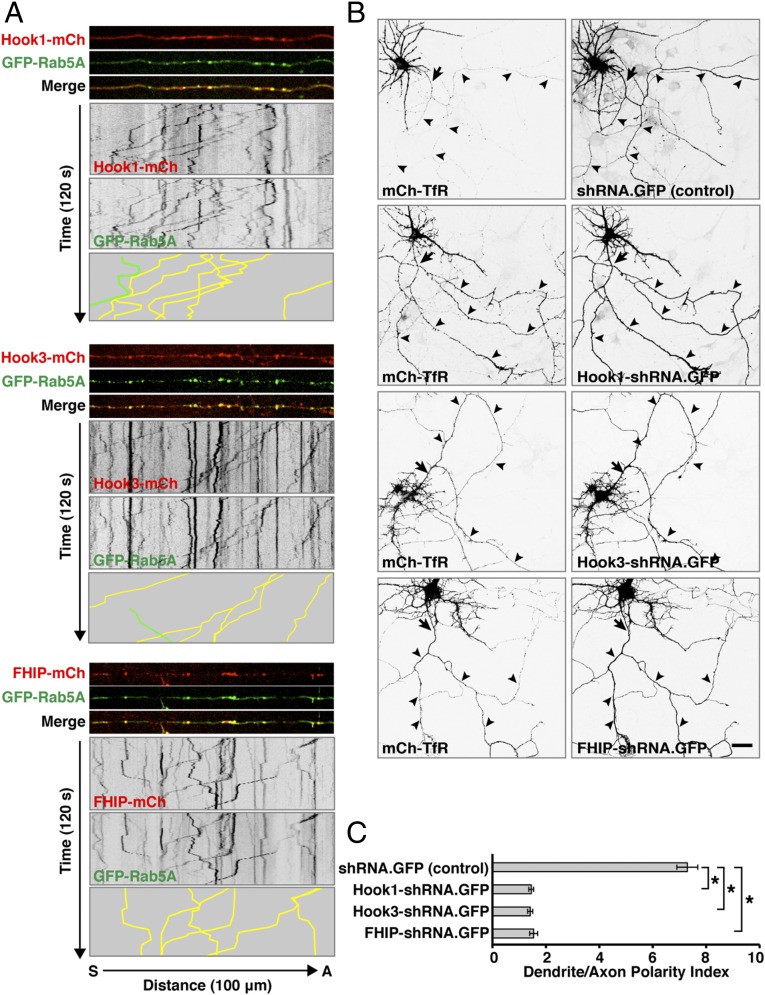
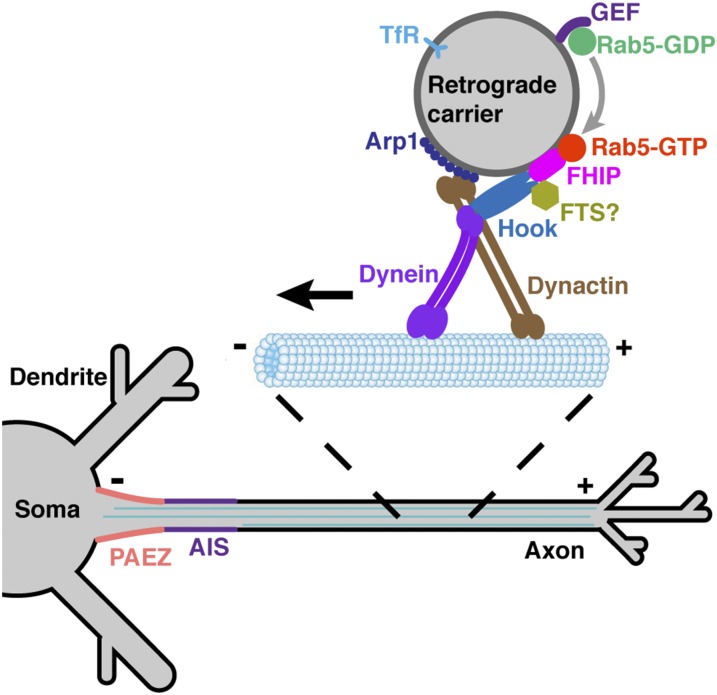
An open question in cell biology is how the general intracellular transport machinery is adapted to perform specialized functions in polarized cells such as neurons. Here we illustrate this adaptation by elucidating a role for the ubiquitous small GTPase Ras-related protein in brain 5 (Rab5) in neuronal polarity. We show that inactivation or depletion of Rab5 in rat hippocampal neurons abrogates the somatodendritic polarity of the transferrin receptor and several glutamate receptor types, resulting in their appearance in the axon. This loss of polarity is not caused primarily by increased transport from the soma to the axon but rather by decreased retrieval from the axon to the soma. Retrieval is also dependent on the Rab5 effector Fused Toes (FTS)-Hook-FTS and Hook-interacting protein (FHIP) (FHF) complex, which interacts with the minus-end-directed microtubule motor dynein and its activator dynactin to drive a population of axonal retrograde carriers containing somatodendritic proteins toward the soma. These findings emphasize the importance of both biosynthetic sorting and axonal retrieval for the polarized distribution of somatodendritic receptors at steady state.
Keywords: FHF complex; Rab5; dynein; neuronal polarity; retrograde transport.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
Cytoplasmic dynein-1 cargo diversity is mediated by the combinatorial assembly of FTS-Hook-FHIP complexes.Elife. 2021 Dec 9;10:e74538. doi: 10.7554/eLife.74538. Elife. 2021. PMID: 34882091 Free PMC article.
-
Dynein activator Hook1 is required for trafficking of BDNF-signaling endosomes in neurons.J Cell Biol. 2019 Jan 7;218(1):220-233. doi: 10.1083/jcb.201805016. Epub 2018 Oct 29. J Cell Biol. 2019. PMID: 30373907 Free PMC article.
-
Retrograde transport of Akt by a neuronal Rab5-APPL1 endosome.Sci Rep. 2019 Feb 21;9(1):2433. doi: 10.1038/s41598-019-38637-0. Sci Rep. 2019. PMID: 30792402 Free PMC article.
-
Cytoplasmic dynein and early endosome transport.Cell Mol Life Sci. 2015 Sep;72(17):3267-80. doi: 10.1007/s00018-015-1926-y. Epub 2015 May 23. Cell Mol Life Sci. 2015. PMID: 26001903 Free PMC article. Review.
-
The function of the axon initial segment in neuronal polarity.Dev Biol. 2022 Sep;489:47-54. doi: 10.1016/j.ydbio.2022.05.016. Epub 2022 May 28. Dev Biol. 2022. PMID: 35640681 Review.
Cited by
-
Analysis of CDPK1 targets identifies a trafficking adaptor complex that regulates microneme exocytosis in Toxoplasma.Elife. 2023 Nov 7;12:RP85654. doi: 10.7554/eLife.85654. Elife. 2023. PMID: 37933960 Free PMC article.
-
Spatial control of membrane traffic in neuronal dendrites.Mol Cell Neurosci. 2020 Jun;105:103492. doi: 10.1016/j.mcn.2020.103492. Epub 2020 Apr 12. Mol Cell Neurosci. 2020. PMID: 32294508 Free PMC article. Review.
-
Temporal Patterns of Angular Displacement of Endosomes: Insights into Motor Protein Exchange Dynamics.Adv Sci (Weinh). 2024 Aug;11(29):e2306849. doi: 10.1002/advs.202306849. Epub 2024 Jun 3. Adv Sci (Weinh). 2024. PMID: 38828676 Free PMC article.
-
Moving and positioning the endolysosomal system.Curr Opin Cell Biol. 2017 Aug;47:1-8. doi: 10.1016/j.ceb.2017.01.008. Epub 2017 Feb 21. Curr Opin Cell Biol. 2017. PMID: 28231489 Free PMC article. Review.
-
Hook3 is a scaffold for the opposite-polarity microtubule-based motors cytoplasmic dynein-1 and KIF1C.J Cell Biol. 2019 Sep 2;218(9):2982-3001. doi: 10.1083/jcb.201812170. Epub 2019 Jul 18. J Cell Biol. 2019. PMID: 31320392 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
