

Plexins function in epithelial repair in both Drosophila and zebrafish
- PMID: 27452696
- PMCID: PMC4962468
- DOI: 10.1038/ncomms12282
Plexins function in epithelial repair in both Drosophila and zebrafish
Abstract
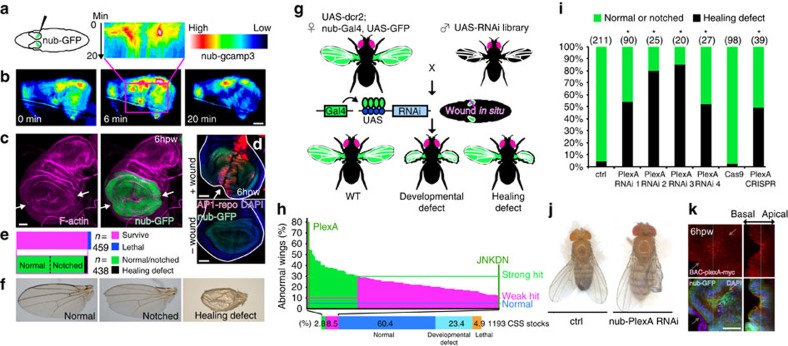
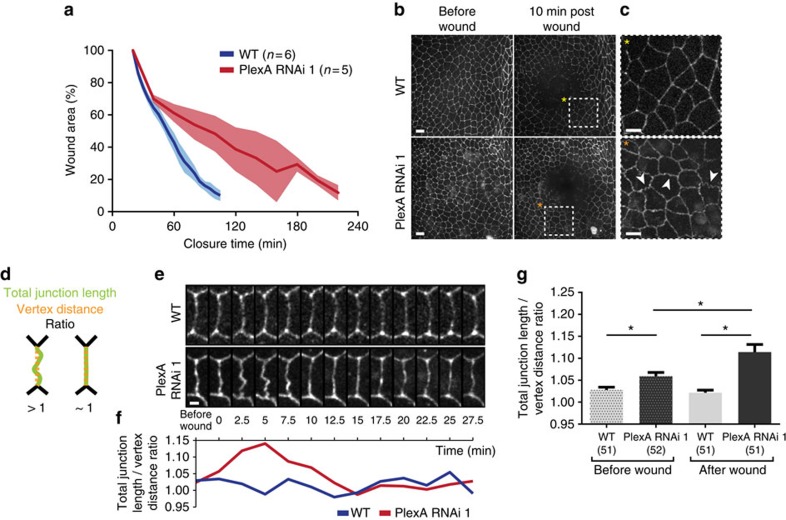
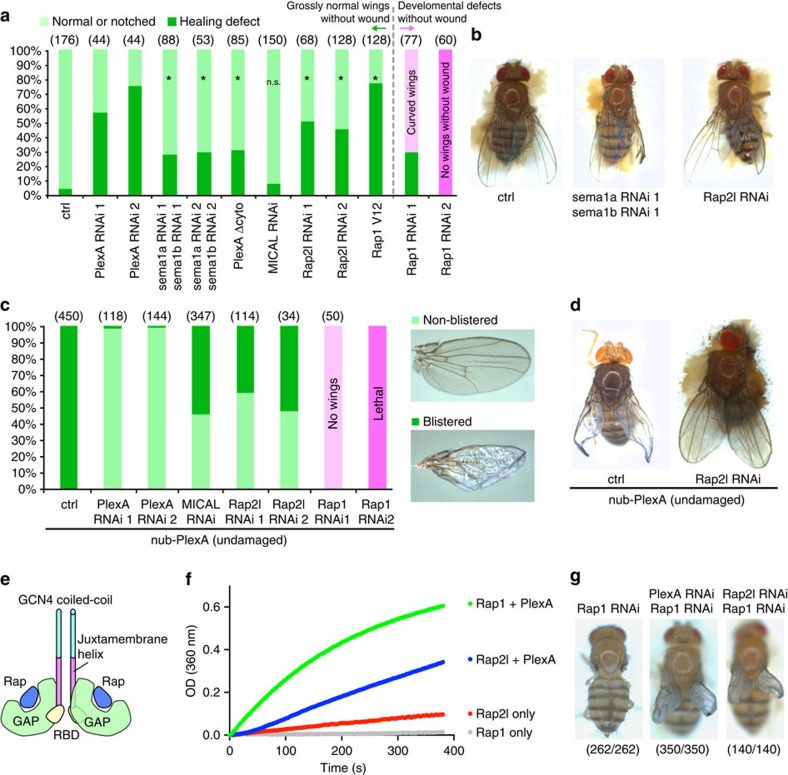
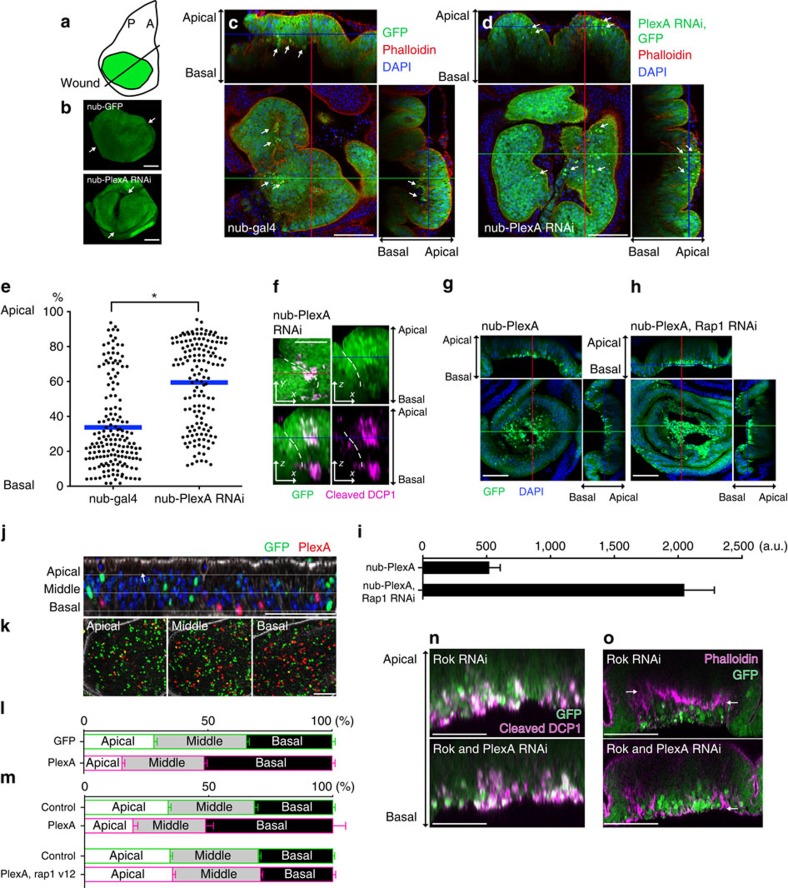
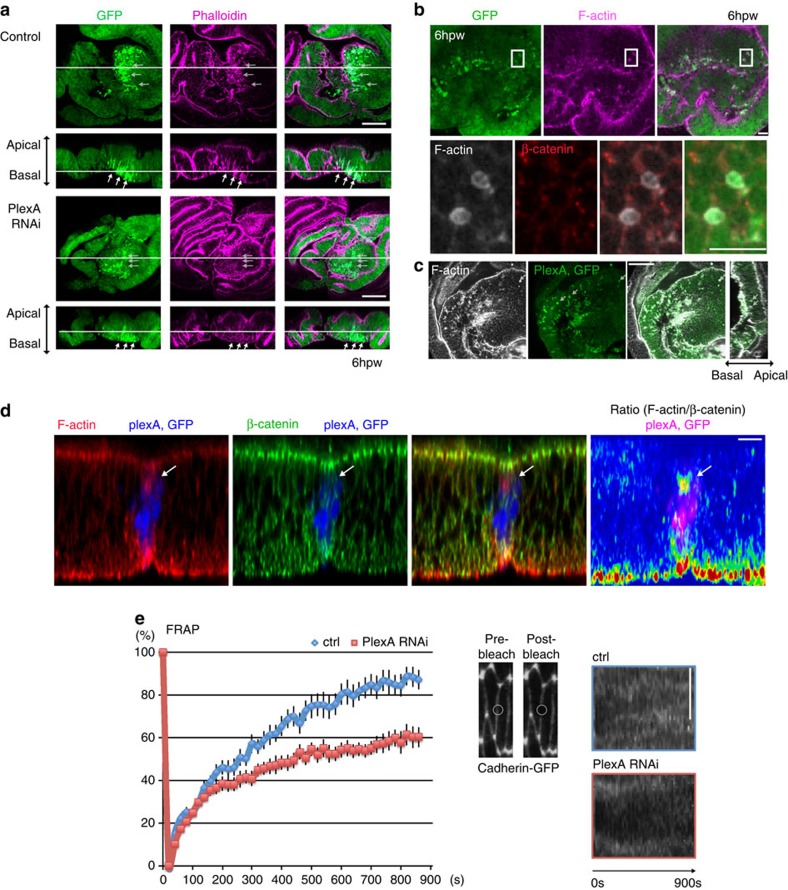
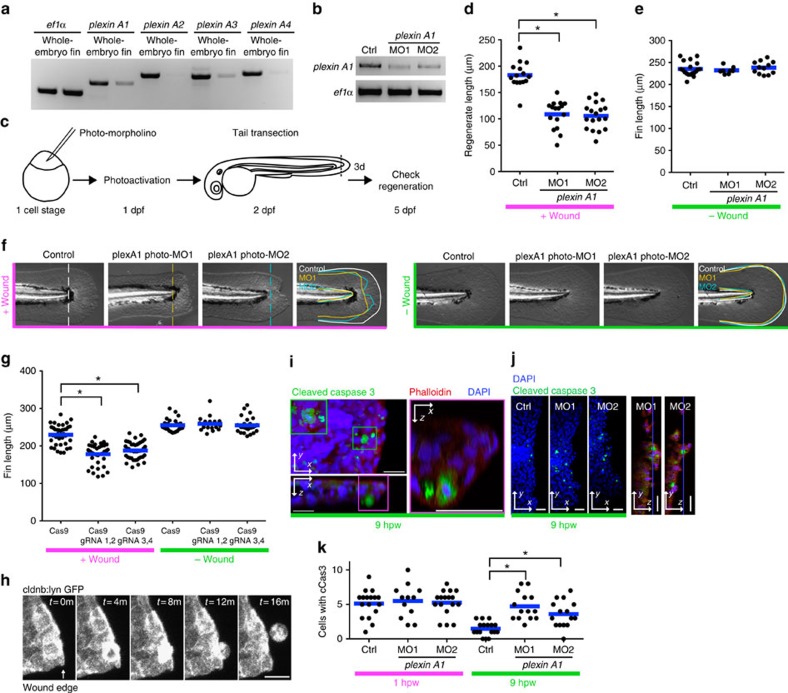
In most multicellular organisms, homeostasis is contingent upon maintaining epithelial integrity. When unanticipated insults breach epithelial barriers, dormant programmes of tissue repair are immediately activated. However, many of the mechanisms that repair damaged epithelia remain poorly characterized. Here we describe a role for Plexin A (PlexA), a protein with particularly well-characterized roles in axonal pathfinding, in the healing of damaged epithelia in Drosophila. Semaphorins, which are PlexA ligands, also regulate tissue repair. We show that Drosophila PlexA has GAP activity for the Rap1 GTPase, which is known to regulate the stability of adherens junctions. Our observations suggest that the inhibition of Rap1 activity by PlexA in damaged Drosophila epithelia allows epithelial remodelling, thus facilitating wound repair. We also demonstrate a role for Plexin A1, a zebrafish orthologue of Drosophila PlexA, in epithelial repair in zebrafish tail fins. Thus, plexins function in epithelial wound healing in diverse taxa.
Figures
Similar articles
-
The axon guidance function of Rap1 small GTPase is independent of PlexA RasGAP activity in Drosophila.Dev Biol. 2016 Oct 15;418(2):258-67. doi: 10.1016/j.ydbio.2016.08.026. Epub 2016 Aug 24. Dev Biol. 2016. PMID: 27565025
-
MICALs, a family of conserved flavoprotein oxidoreductases, function in plexin-mediated axonal repulsion.Cell. 2002 Jun 28;109(7):887-900. doi: 10.1016/s0092-8674(02)00794-8. Cell. 2002. PMID: 12110185
-
Semaphorin-Plexin Signaling Controls Mitotic Spindle Orientation during Epithelial Morphogenesis and Repair.Dev Cell. 2015 May 4;33(3):299-313. doi: 10.1016/j.devcel.2015.02.001. Epub 2015 Apr 16. Dev Cell. 2015. PMID: 25892012
-
Tension (re)builds: Biophysical mechanisms of embryonic wound repair.Mech Dev. 2017 Apr;144(Pt A):43-52. doi: 10.1016/j.mod.2016.11.004. Epub 2016 Dec 15. Mech Dev. 2017. PMID: 27989746 Review.
-
Connexin Communication Compartments and Wound Repair in Epithelial Tissue.Int J Mol Sci. 2018 May 3;19(5):1354. doi: 10.3390/ijms19051354. Int J Mol Sci. 2018. PMID: 29751558 Free PMC article. Review.
Cited by
-
JNK-mediated Slit-Robo signaling facilitates epithelial wound repair by extruding dying cells.Sci Rep. 2019 Dec 20;9(1):19549. doi: 10.1038/s41598-019-56137-z. Sci Rep. 2019. PMID: 31863086 Free PMC article.
-
Kynurenine Metabolism in the Fat Body Non-autonomously Regulates Imaginal Disc Repair in Drosophila.iScience. 2020 Nov 6;23(12):101738. doi: 10.1016/j.isci.2020.101738. eCollection 2020 Dec 18. iScience. 2020. PMID: 33376969 Free PMC article.
-
Single-cell transcriptomes of mouse bladder urothelium uncover novel cell type markers and urothelial differentiation characteristics.Cell Prolif. 2021 Apr;54(4):e13007. doi: 10.1111/cpr.13007. Epub 2021 Feb 3. Cell Prolif. 2021. PMID: 33538002 Free PMC article.
-
Erebosis, a new cell death mechanism during homeostatic turnover of gut enterocytes.PLoS Biol. 2022 Apr 25;20(4):e3001586. doi: 10.1371/journal.pbio.3001586. eCollection 2022 Apr. PLoS Biol. 2022. PMID: 35468130 Free PMC article.
-
Model systems for regeneration: Drosophila.Development. 2020 Apr 6;147(7):dev173781. doi: 10.1242/dev.173781. Development. 2020. PMID: 32253254 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
