

DNA minicircles clarify the specific role of DNA structure on retroviral integration
- PMID: 27439712
- PMCID: PMC5027509
- DOI: 10.1093/nar/gkw651
DNA minicircles clarify the specific role of DNA structure on retroviral integration
Abstract
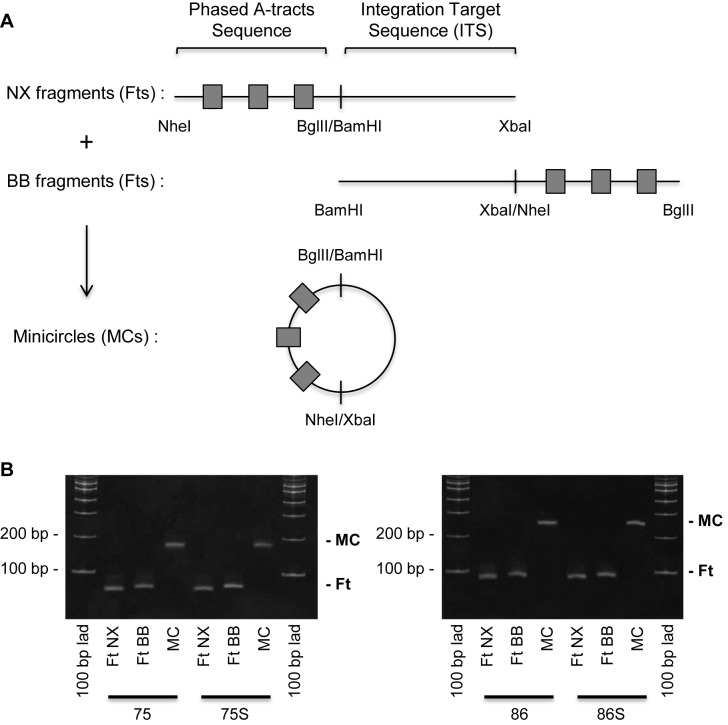
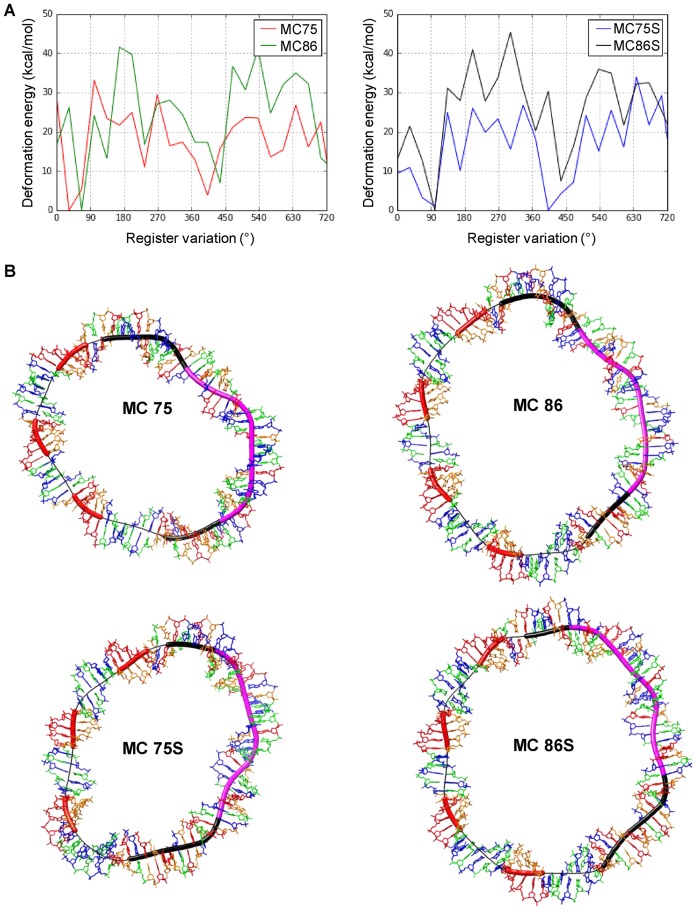
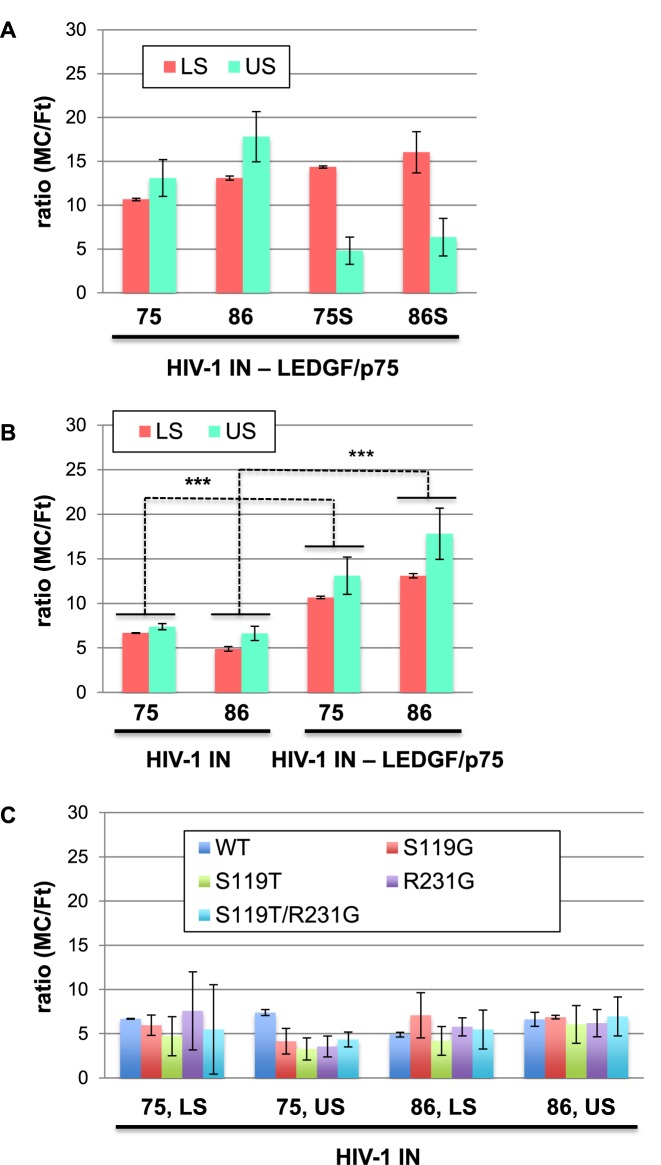
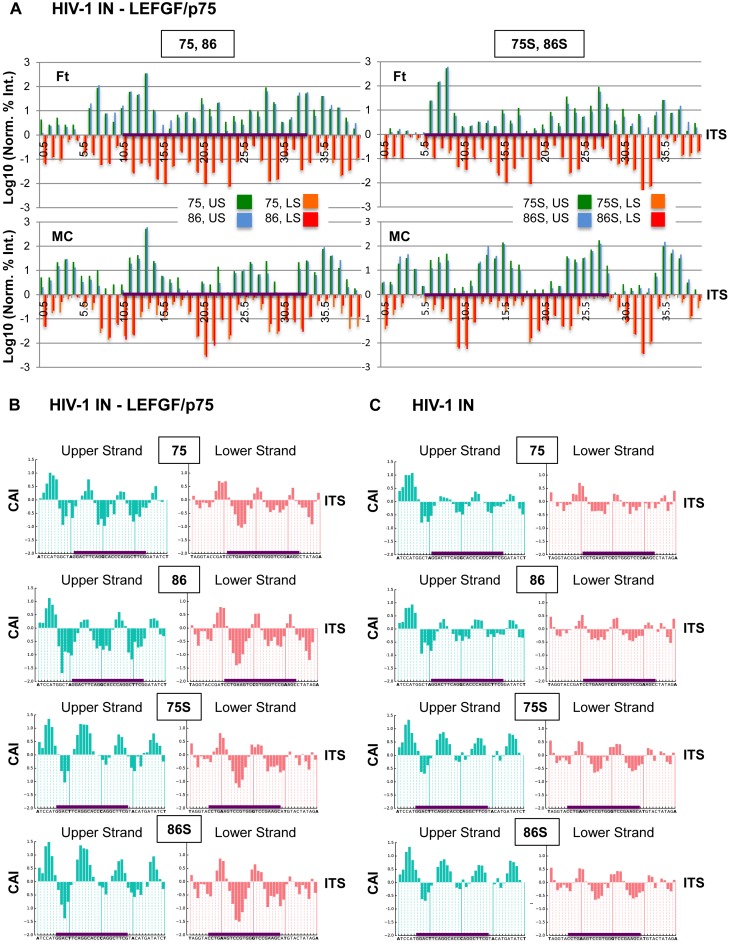
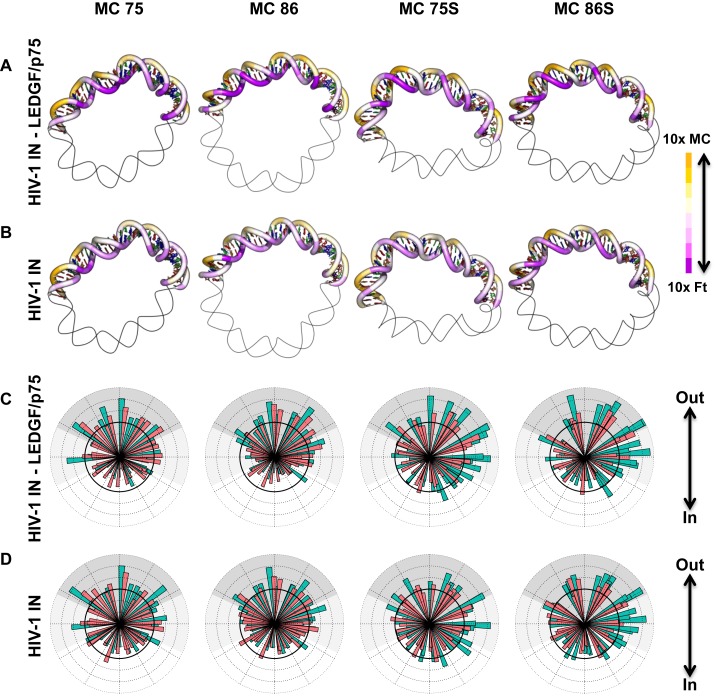
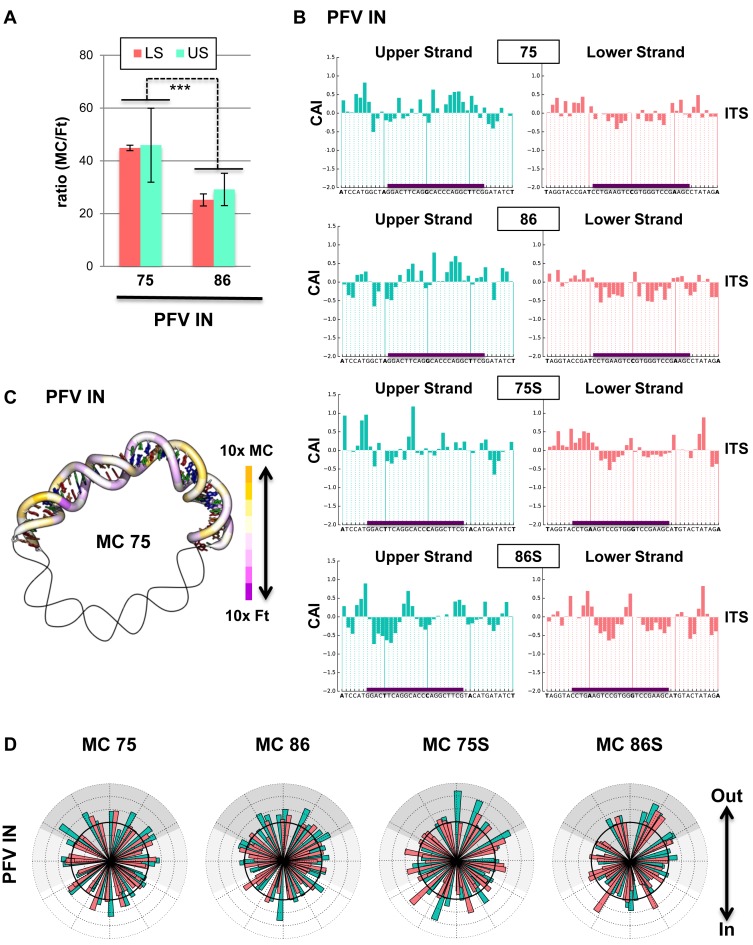
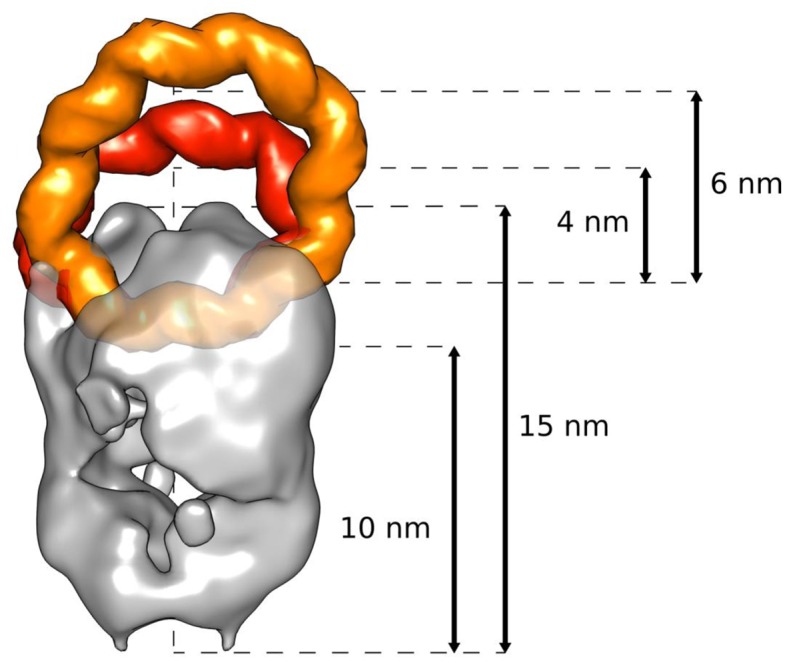
Chromatin regulates the selectivity of retroviral integration into the genome of infected cells. At the nucleosome level, both histones and DNA structure are involved in this regulation. We propose a strategy that allows to specifically study a single factor: the DNA distortion induced by the nucleosome. This strategy relies on mimicking this distortion using DNA minicircles (MCs) having a fixed rotational orientation of DNA curvature, coupled with atomic-resolution modeling. Contrasting MCs with linear DNA fragments having identical sequences enabled us to analyze the impact of DNA distortion on the efficiency and selectivity of integration. We observed a global enhancement of HIV-1 integration in MCs and an enrichment of integration sites in the outward-facing DNA major grooves. Both of these changes are favored by LEDGF/p75, revealing a new, histone-independent role of this integration cofactor. PFV integration is also enhanced in MCs, but is not associated with a periodic redistribution of integration sites, thus highlighting its distinct catalytic properties. MCs help to separate the roles of target DNA structure, histone modifications and integrase (IN) cofactors during retroviral integration and to reveal IN-specific regulation mechanisms.
© The Author(s) 2016. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
Modulation of chromatin structure by the FACT histone chaperone complex regulates HIV-1 integration.Retrovirology. 2017 Jul 28;14(1):39. doi: 10.1186/s12977-017-0363-4. Retrovirology. 2017. PMID: 28754126 Free PMC article.
-
Chromatinized templates reveal the requirement for the LEDGF/p75 PWWP domain during HIV-1 integration in vitro.Nucleic Acids Res. 2008 Mar;36(4):1237-46. doi: 10.1093/nar/gkm1127. Epub 2008 Jan 3. Nucleic Acids Res. 2008. PMID: 18174227 Free PMC article.
-
Intasome architecture and chromatin density modulate retroviral integration into nucleosome.Retrovirology. 2015 Feb 7;12:13. doi: 10.1186/s12977-015-0145-9. Retrovirology. 2015. PMID: 25807893 Free PMC article.
-
[LEDGF/p75: a novel target for anti-HIV therapy and advances in the study of its related inhibitors].Yao Xue Xue Bao. 2009 Sep;44(9):953-60. Yao Xue Xue Bao. 2009. PMID: 20055168 Review. Chinese.
-
HIV DNA integration.Cold Spring Harb Perspect Med. 2012 Jul;2(7):a006890. doi: 10.1101/cshperspect.a006890. Cold Spring Harb Perspect Med. 2012. PMID: 22762018 Free PMC article. Review.
Cited by
-
Structural interplay between DNA-shape protein recognition and supercoiling: The case of IHF.Comput Struct Biotechnol J. 2022 Sep 19;20:5264-5274. doi: 10.1016/j.csbj.2022.09.020. eCollection 2022. Comput Struct Biotechnol J. 2022. PMID: 36212531 Free PMC article.
-
Human H4 tail stimulates HIV-1 integration through binding to the carboxy-terminal domain of integrase.Nucleic Acids Res. 2019 Apr 23;47(7):3607-3618. doi: 10.1093/nar/gkz091. Nucleic Acids Res. 2019. PMID: 30767014 Free PMC article.
-
Integrating transposable elements in the 3D genome.Mob DNA. 2020 Feb 4;11:8. doi: 10.1186/s13100-020-0202-3. eCollection 2020. Mob DNA. 2020. PMID: 32042316 Free PMC article.
-
Retroviral integrase protein and intasome nucleoprotein complex structures.World J Biol Chem. 2017 Feb 26;8(1):32-44. doi: 10.4331/wjbc.v8.i1.32. World J Biol Chem. 2017. PMID: 28289517 Free PMC article. Review.
-
Modulation of the functional association between the HIV-1 intasome and the nucleosome by histone amino-terminal tails.Retrovirology. 2017 Nov 28;14(1):54. doi: 10.1186/s12977-017-0378-x. Retrovirology. 2017. PMID: 29179726 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
