

Active Control of Repetitive Structural Transitions between Replication Forks and Holliday Junctions by Werner Syndrome Helicase
- PMID: 27427477
- PMCID: PMC5167498
- DOI: 10.1016/j.str.2016.06.004
Active Control of Repetitive Structural Transitions between Replication Forks and Holliday Junctions by Werner Syndrome Helicase
Abstract
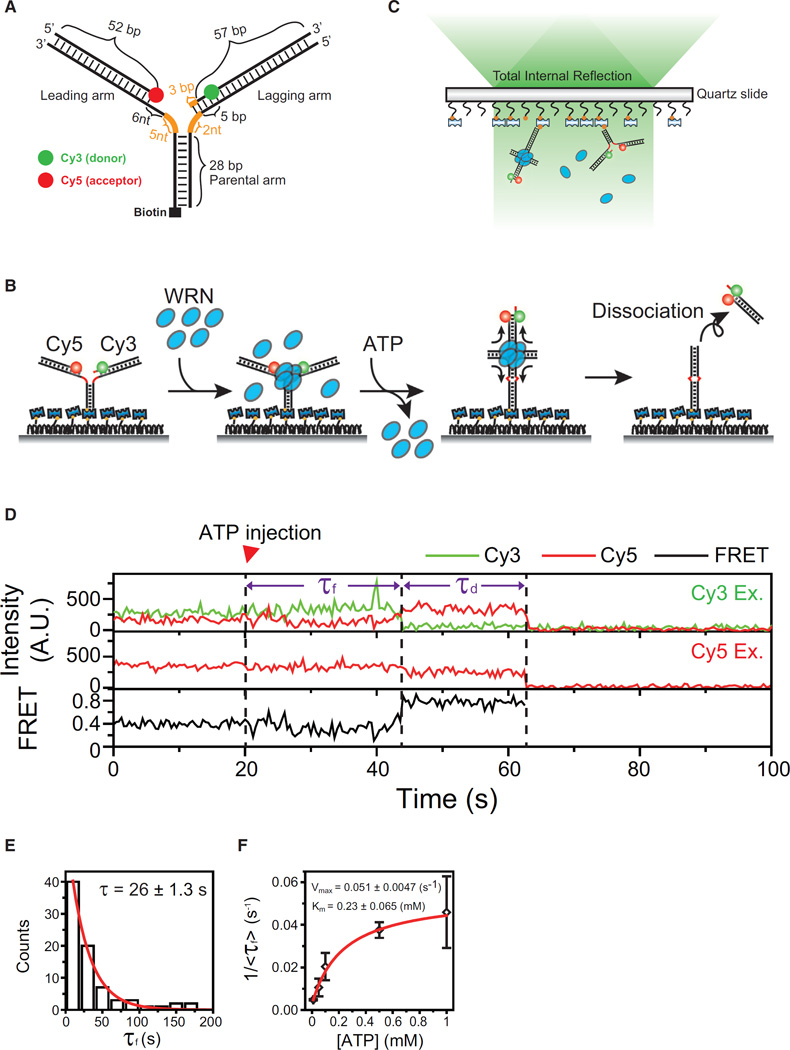
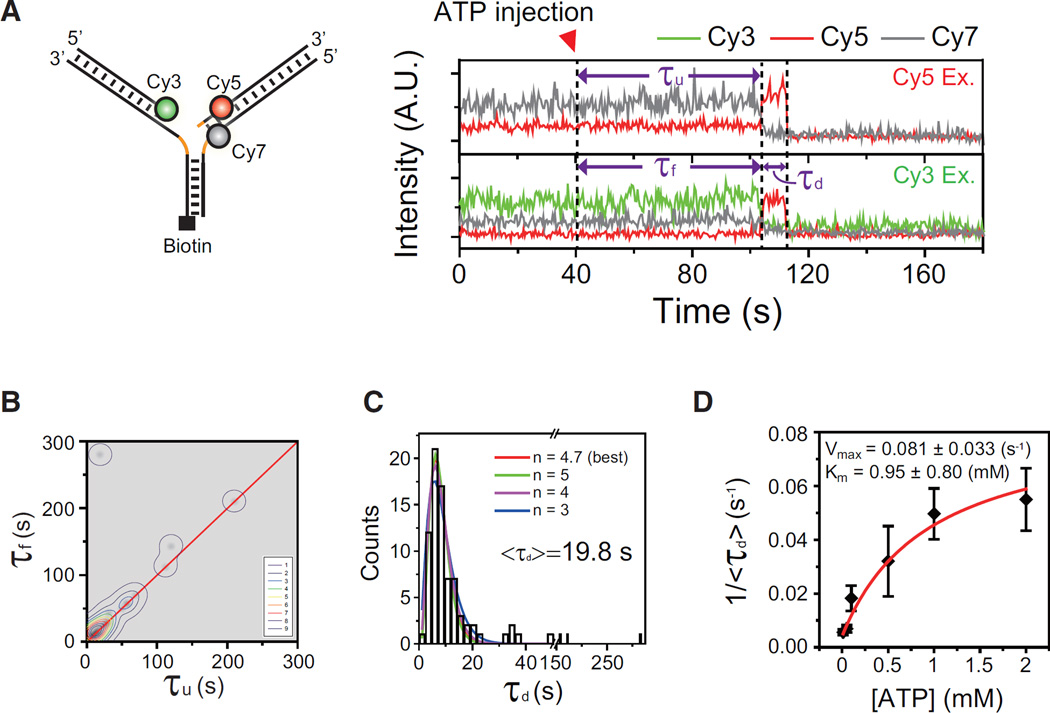
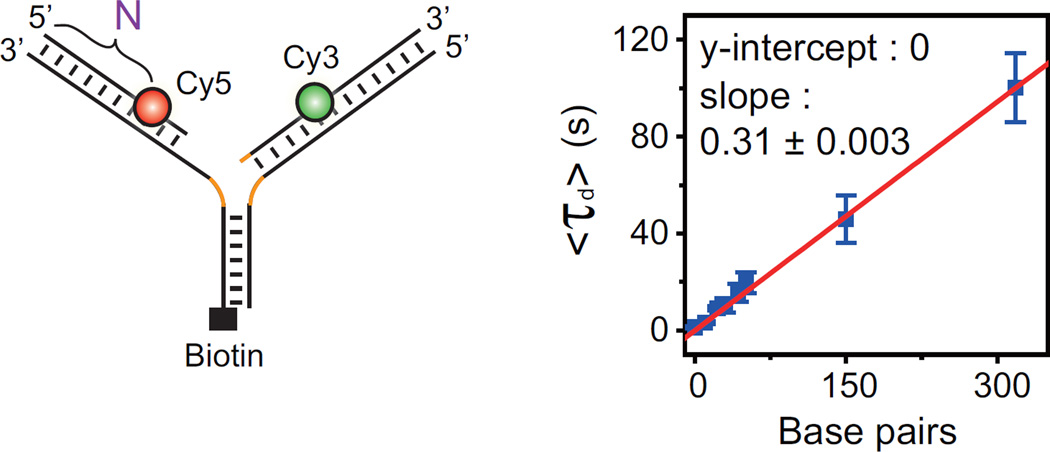
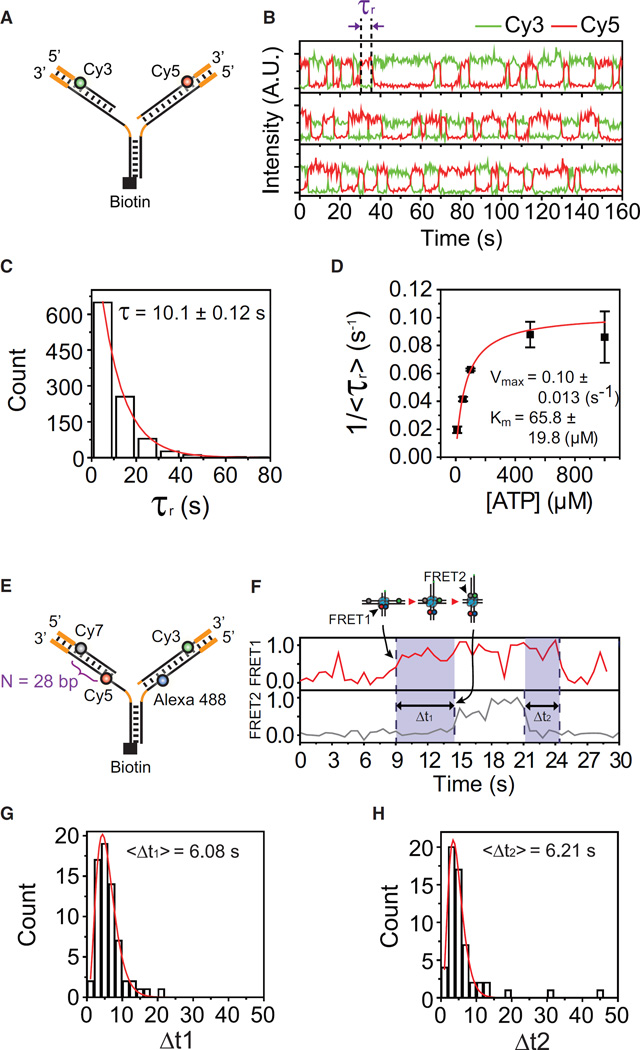
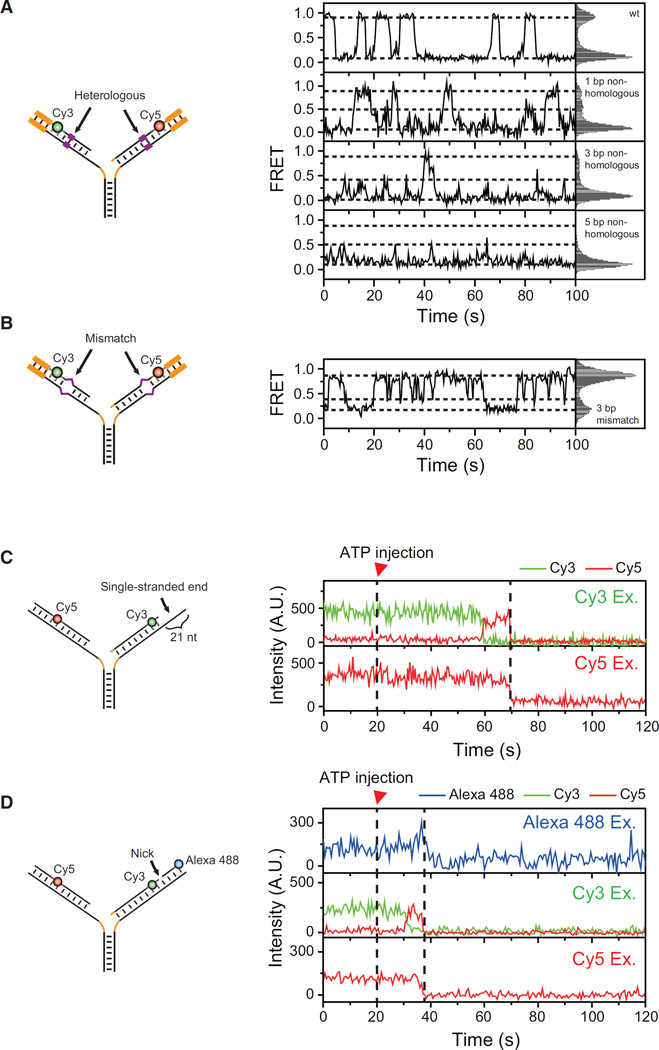
The reactivation of stalled DNA replication via fork regression invokes Holliday junction formation, branch migration, and the recovery of the replication fork after DNA repair or error-free DNA synthesis. The coordination mechanism for these DNA structural transitions by molecular motors, however, remains unclear. Here we perform single-molecule fluorescence experiments with Werner syndrome protein (WRN) and model replication forks. The Holliday junction is readily formed once the lagging arm is unwound, and migrated unidirectionally with 3.2 ± 0.03 bases/s velocity. The recovery of the replication fork was controlled by branch migration reversal of WRN, resulting in repetitive fork regression. The Holliday junction formation, branch migration, and migration direction reversal are all ATP dependent, revealing that WRN uses the energy of ATP hydrolysis to actively coordinate the structural transitions of DNA.
Keywords: replication fork regression; single-molecule FRET; werner syndrome helicase.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Replication fork regression in vitro by the Werner syndrome protein (WRN): holliday junction formation, the effect of leading arm structure and a potential role for WRN exonuclease activity.Nucleic Acids Res. 2007;35(17):5729-47. doi: 10.1093/nar/gkm561. Epub 2007 Aug 23. Nucleic Acids Res. 2007. PMID: 17717003 Free PMC article.
-
The Werner syndrome protein binds replication fork and holliday junction DNAs as an oligomer.J Biol Chem. 2008 Sep 5;283(36):24478-83. doi: 10.1074/jbc.M803370200. Epub 2008 Jul 2. J Biol Chem. 2008. PMID: 18596042 Free PMC article.
-
The Werner and Bloom syndrome proteins help resolve replication blockage by converting (regressed) holliday junctions to functional replication forks.Biochemistry. 2011 Aug 16;50(32):6774-88. doi: 10.1021/bi2001054. Epub 2011 Jul 21. Biochemistry. 2011. PMID: 21736299 Free PMC article.
-
Werner Syndrome Protein and DNA Replication.Int J Mol Sci. 2018 Nov 2;19(11):3442. doi: 10.3390/ijms19113442. Int J Mol Sci. 2018. PMID: 30400178 Free PMC article. Review.
-
Interplay between DNA replication, recombination and repair based on the structure of RecG helicase.Philos Trans R Soc Lond B Biol Sci. 2004 Jan 29;359(1441):49-59. doi: 10.1098/rstb.2003.1364. Philos Trans R Soc Lond B Biol Sci. 2004. PMID: 15065656 Free PMC article. Review.
Cited by
-
Werner syndrome protein works as a dimer for unwinding and replication fork regression.Nucleic Acids Res. 2023 Jan 11;51(1):337-348. doi: 10.1093/nar/gkac1200. Nucleic Acids Res. 2023. PMID: 36583333 Free PMC article.
-
ATAD5 promotes replication restart by regulating RAD51 and PCNA in response to replication stress.Nat Commun. 2019 Dec 16;10(1):5718. doi: 10.1038/s41467-019-13667-4. Nat Commun. 2019. PMID: 31844045 Free PMC article.
-
Genomic instability and DNA replication defects in progeroid syndromes.Nucleus. 2018 Dec 31;9(1):368-379. doi: 10.1080/19491034.2018.1476793. Epub 2018 Jun 23. Nucleus. 2018. PMID: 29936894 Free PMC article. Review.
-
Replication fork regression and its regulation.FEMS Yeast Res. 2017 Jan 1;17(1):fow110. doi: 10.1093/femsyr/fow110. FEMS Yeast Res. 2017. PMID: 28011905 Free PMC article. Review.
References
-
- Biswas I, Yamamoto A, Hsieh P. Branch migration through DNA sequence heterology. J. Mol. Biol. 1998;279:795–806. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
