

Roles of d-Amino Acids on the Bioactivity of Host Defense Peptides
- PMID: 27376281
- PMCID: PMC4964399
- DOI: 10.3390/ijms17071023
Roles of d-Amino Acids on the Bioactivity of Host Defense Peptides
Abstract
Host defense peptides (HDPs) are positively-charged and amphipathic components of the innate immune system that have demonstrated great potential to become the next generation of broad spectrum therapeutic agents effective against a vast array of pathogens and tumor. As such, many approaches have been taken to improve the therapeutic efficacy of HDPs. Amongst these methods, the incorporation of d-amino acids (d-AA) is an approach that has demonstrated consistent success in improving HDPs. Although, virtually all HDP review articles briefly mentioned about the role of d-AA, however it is rather surprising that no systematic review specifically dedicated to this topic exists. Given the impact that d-AA incorporation has on HDPs, this review aims to fill that void with a systematic discussion of the impact of d-AA on HDPs.
Keywords: ">d-amino acid; AMP; HDP; anticancer peptide; antimicrobial peptide; bioactivity; diastereomer; host defense peptide.
Figures
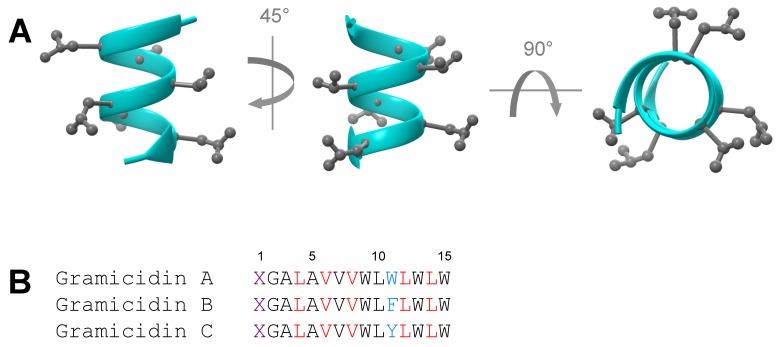
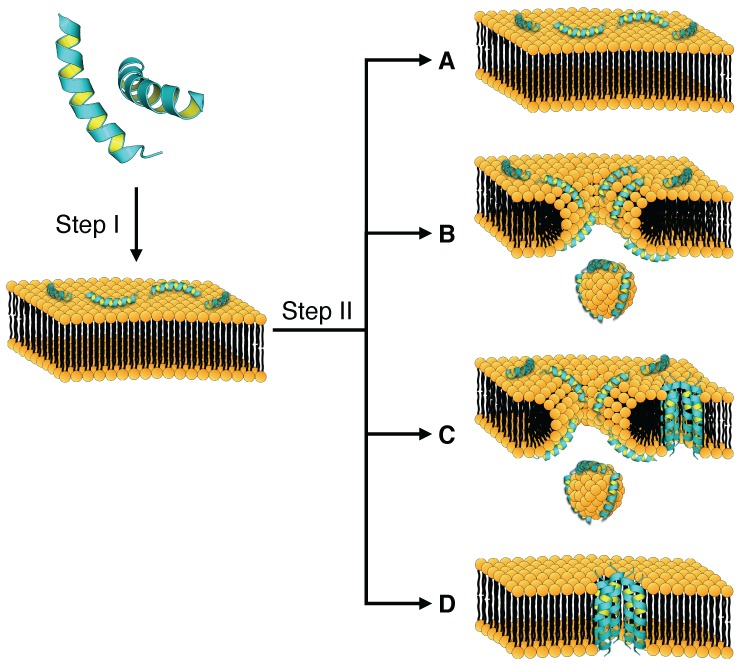
Similar articles
-
Peptide antimicrobials: cell wall as a bacterial target.Ann N Y Acad Sci. 2013 Jan;1277:127-38. doi: 10.1111/nyas.12005. Epub 2013 Jan 9. Ann N Y Acad Sci. 2013. PMID: 23302022 Review.
-
Host Defense Peptides: Dual Antimicrobial and Immunomodulatory Action.Int J Mol Sci. 2021 Oct 16;22(20):11172. doi: 10.3390/ijms222011172. Int J Mol Sci. 2021. PMID: 34681833 Free PMC article. Review.
-
Structural determinants of host defense peptides for antimicrobial activity and target cell selectivity.Biochimie. 2010 Sep;92(9):1236-41. doi: 10.1016/j.biochi.2010.02.023. Epub 2010 Feb 25. Biochimie. 2010. PMID: 20188791 Review.
-
Deletion of all cysteines in tachyplesin I abolishes hemolytic activity and retains antimicrobial activity and lipopolysaccharide selective binding.Biochemistry. 2006 May 23;45(20):6529-40. doi: 10.1021/bi052629q. Biochemistry. 2006. PMID: 16700563 Free PMC article.
-
Effect of stereochemistry, chain length and sequence pattern on antimicrobial properties of short synthetic β-sheet forming peptide amphiphiles.Biomaterials. 2014 Jan;35(4):1315-25. doi: 10.1016/j.biomaterials.2013.10.053. Epub 2013 Nov 7. Biomaterials. 2014. PMID: 24211081
Cited by
-
Toward insights on determining factors for high activity in antimicrobial peptides via machine learning.PeerJ. 2019 Dec 20;7:e8265. doi: 10.7717/peerj.8265. eCollection 2019. PeerJ. 2019. PMID: 31875156 Free PMC article.
-
D1018 with higher stability and excellent lipopolysaccharide binding affinity has potent anti-bacterial and anti-inflammatory activity.Front Microbiol. 2022 Dec 1;13:1010017. doi: 10.3389/fmicb.2022.1010017. eCollection 2022. Front Microbiol. 2022. PMID: 36532445 Free PMC article.
-
Antimicrobial Peptides and Their Therapeutic Potential for Bacterial Skin Infections and Wounds.Front Pharmacol. 2018 Mar 28;9:281. doi: 10.3389/fphar.2018.00281. eCollection 2018. Front Pharmacol. 2018. PMID: 29643807 Free PMC article. Review.
-
D-form KLKLLLLLKLK-NH2 peptide exerts higher antimicrobial properties than its L-form counterpart via an association with bacterial cell wall components.Sci Rep. 2017 Mar 6;7:43384. doi: 10.1038/srep43384. Sci Rep. 2017. PMID: 28262682 Free PMC article.
-
Antimicrobial Activity of Small Synthetic Peptides Based on the Marine Peptide Turgencin A: Prediction of Antimicrobial Peptide Sequences in a Natural Peptide and Strategy for Optimization of Potency.Int J Mol Sci. 2020 Jul 30;21(15):5460. doi: 10.3390/ijms21155460. Int J Mol Sci. 2020. PMID: 32751755 Free PMC article.
References
-
- Lynn M.A., Kindrachuk J., Marr A.K., Jenssen H., Pante N., Elliott M.R., Napper S., Hancock R.E., McMaster W.R. Effect of BMAP-28 antimicrobial peptides on Leishmania major promastigote and amastigote growth: Role of leishmanolysin in parasite survival. PLoS Negl. Trop. Dis. 2011;5:1–13. doi: 10.1371/journal.pntd.0001141. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
