

Mutation in the Pro-Peptide Region of a Cysteine Protease Leads to Altered Activity and Specificity-A Structural and Biochemical Approach
- PMID: 27352302
- PMCID: PMC4924875
- DOI: 10.1371/journal.pone.0158024
Mutation in the Pro-Peptide Region of a Cysteine Protease Leads to Altered Activity and Specificity-A Structural and Biochemical Approach
Abstract
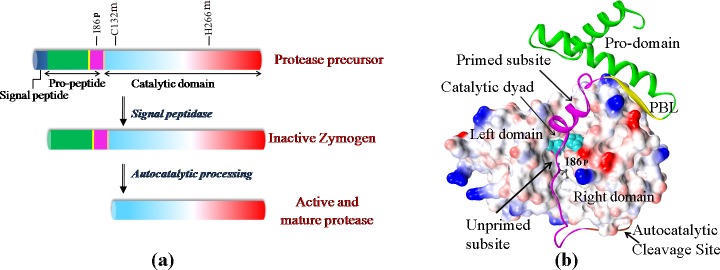
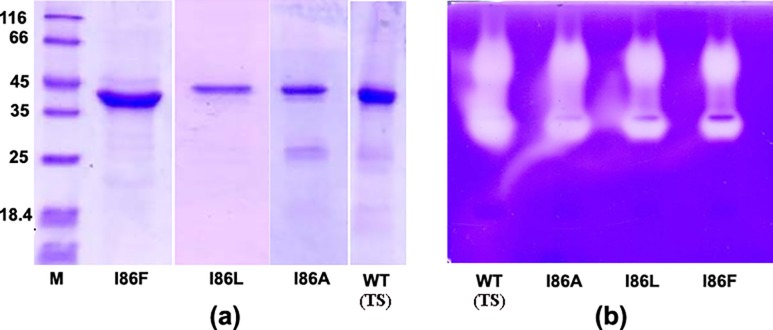
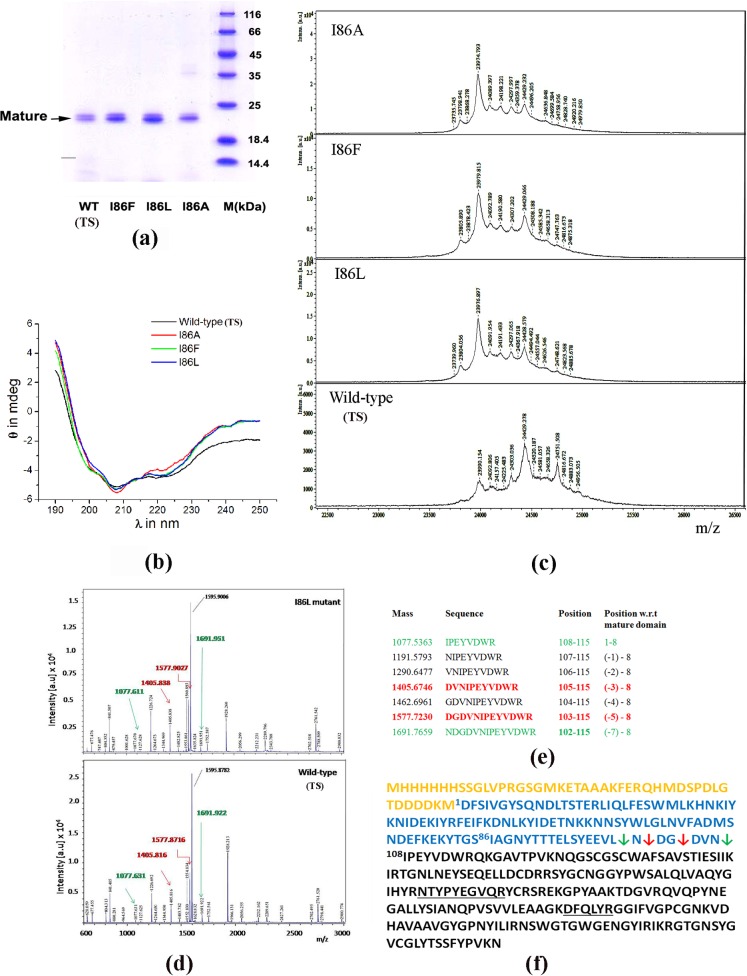
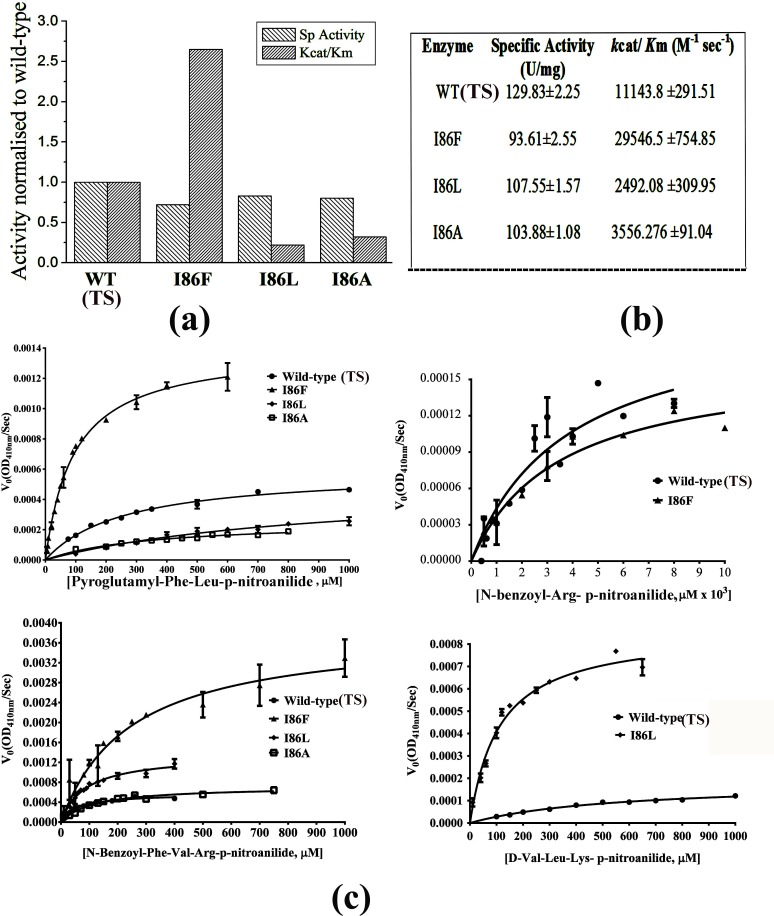
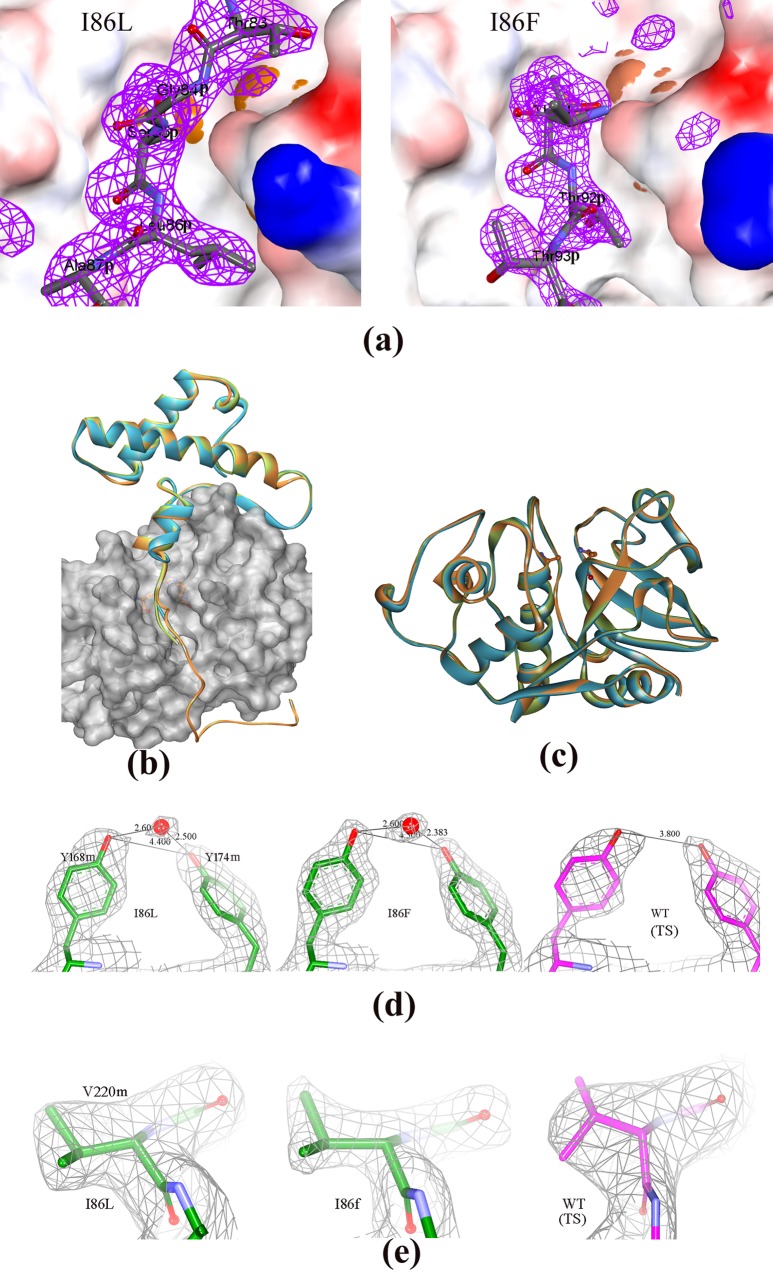
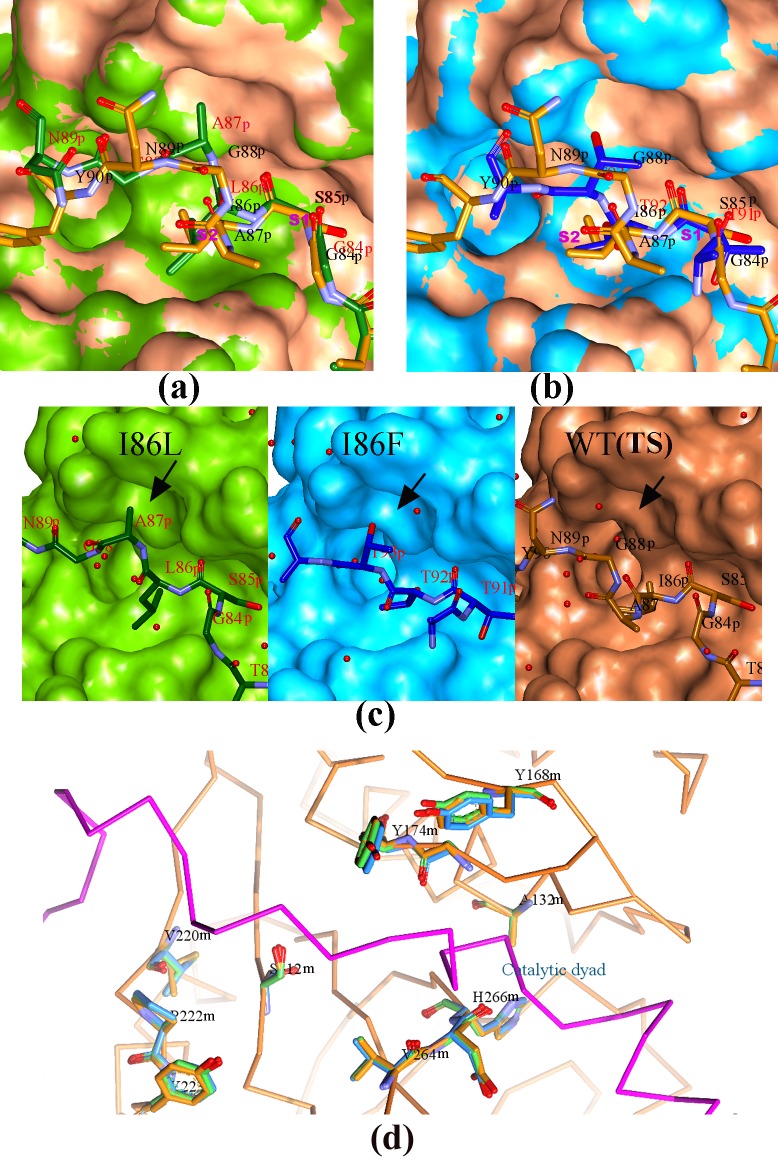
Papain-like proteases contain an N-terminal pro-peptide in their zymogen form that is important for correct folding and spatio-temporal regulation of the proteolytic activity of these proteases. Catalytic removal of the pro-peptide is required for the protease to become active. In this study, we have generated three different mutants of papain (I86F, I86L and I86A) by replacing the residue I86 in its pro-peptide region, which blocks the specificity determining S2-subsite of the catalytic cleft of the protease in its zymogen form with a view to investigate the effect of mutation on the catalytic activity of the protease. Steady-state enzyme kinetic analyses of the corresponding mutant proteases with specific peptide substrates show significant alteration of substrate specificity-I86F and I86L have 2.7 and 29.1 times higher kcat/Km values compared to the wild-type against substrates having Phe and Leu at P2 position, respectively, while I86A shows lower catalytic activity against majority of the substrates tested. Far-UV CD scan and molecular mass analyses of the mature form of the mutant proteases reveal similar CD spectra and intact masses to that of the wild-type. Crystal structures of zymogens of I86F and I86L mutants suggest that subtle reorganization of active site residues, including water, upon binding of the pro-peptide may allow the enzyme to achieve discriminatory substrate selectivity and catalytic efficiency. However, accurate and reliable predictions on alteration of substrate specificity require atomic resolution structure of the catalytic domain after zymogen activation, which remains a challenging task. In this study we demonstrate that through single amino acid substitution in pro-peptide, it is possible to modify the substrate specificity of papain and hence the pro-peptide of a protease can also be a useful target for altering its catalytic activity/specificity.
Conflict of interest statement
Figures
Similar articles
-
The structure of a thermostable mutant of pro-papain reveals its activation mechanism.Acta Crystallogr D Biol Crystallogr. 2012 Dec;68(Pt 12):1591-603. doi: 10.1107/S0907444912038607. Epub 2012 Nov 9. Acta Crystallogr D Biol Crystallogr. 2012. PMID: 23151624
-
Processing of the papain precursor. The ionization state of a conserved amino acid motif within the Pro region participates in the regulation of intramolecular processing.J Biol Chem. 1995 May 5;270(18):10838-46. doi: 10.1074/jbc.270.18.10838. J Biol Chem. 1995. PMID: 7738022
-
Enhancement of proteolytic activity of a thermostable papain-like protease by structure-based rational design.PLoS One. 2013 May 3;8(5):e62619. doi: 10.1371/journal.pone.0062619. Print 2013. PLoS One. 2013. PMID: 23671614 Free PMC article.
-
Protease propeptide structures, mechanisms of activation, and functions.Crit Rev Biochem Mol Biol. 2020 Apr;55(2):111-165. doi: 10.1080/10409238.2020.1742090. Epub 2020 Apr 14. Crit Rev Biochem Mol Biol. 2020. PMID: 32290726 Review.
-
Proregion structure of members of the papain superfamily. Mode of inhibition of enzymatic activity.Biochimie. 1997 Nov;79(11):645-52. doi: 10.1016/s0300-9084(97)83497-9. Biochimie. 1997. PMID: 9479446 Review.
Cited by
-
Nature-inspired Enzyme engineering and sustainable catalysis: biochemical clues from the world of plants and extremophiles.Front Bioeng Biotechnol. 2023 Jun 20;11:1229300. doi: 10.3389/fbioe.2023.1229300. eCollection 2023. Front Bioeng Biotechnol. 2023. PMID: 37409164 Free PMC article. Review.
-
Characterization of a novel cathepsin L-like protease from Taenia solium metacestodes for the immunodiagnosis of porcine cysticercosis.Vet Parasitol. 2019 Mar;267:9-16. doi: 10.1016/j.vetpar.2019.01.004. Epub 2019 Jan 29. Vet Parasitol. 2019. PMID: 30878092 Free PMC article.
-
Structure determinants defining the specificity of papain-like cysteine proteases.Comput Struct Biotechnol J. 2022 Nov 24;20:6552-6569. doi: 10.1016/j.csbj.2022.11.040. eCollection 2022. Comput Struct Biotechnol J. 2022. PMID: 36467578 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
