

Structural Elements in the Gαs and Gαq C Termini That Mediate Selective G Protein-coupled Receptor (GPCR) Signaling
- PMID: 27330078
- PMCID: PMC5016181
- DOI: 10.1074/jbc.M116.735720
Structural Elements in the Gαs and Gαq C Termini That Mediate Selective G Protein-coupled Receptor (GPCR) Signaling
Abstract
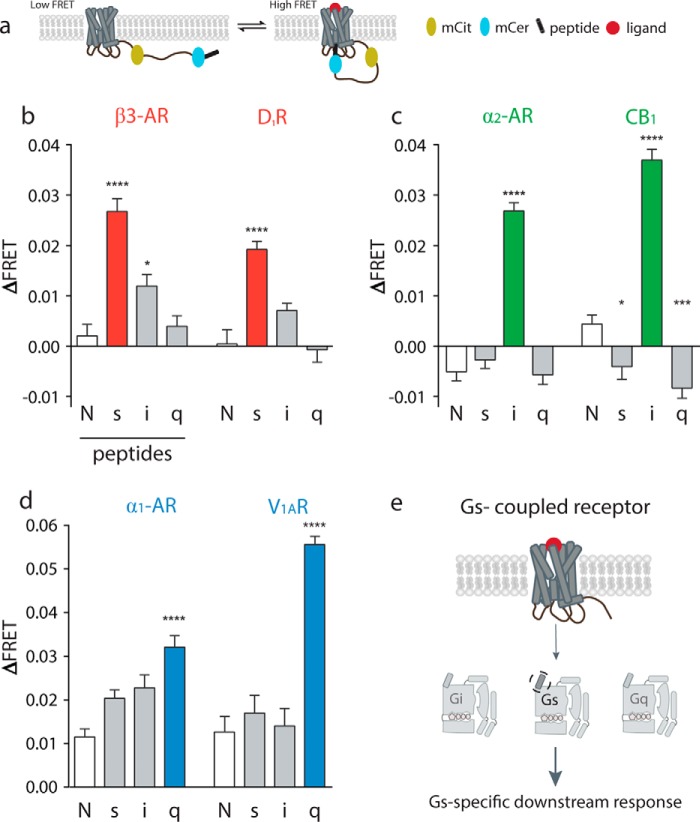
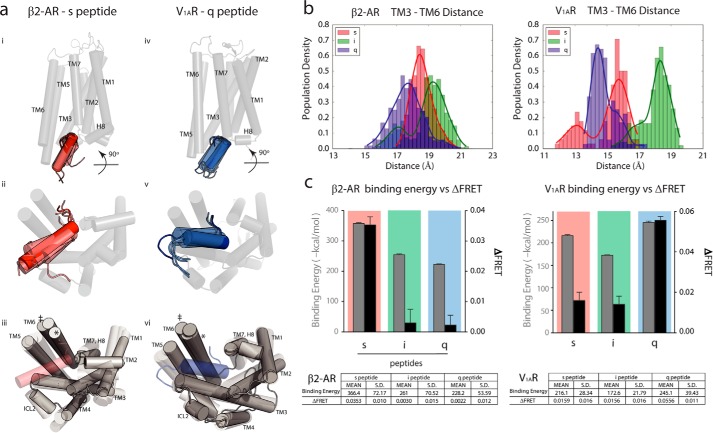
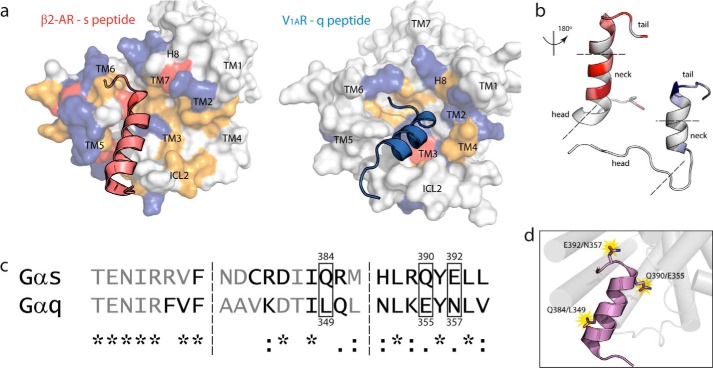
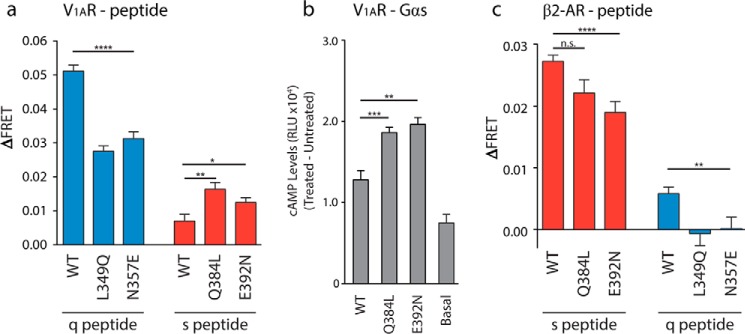
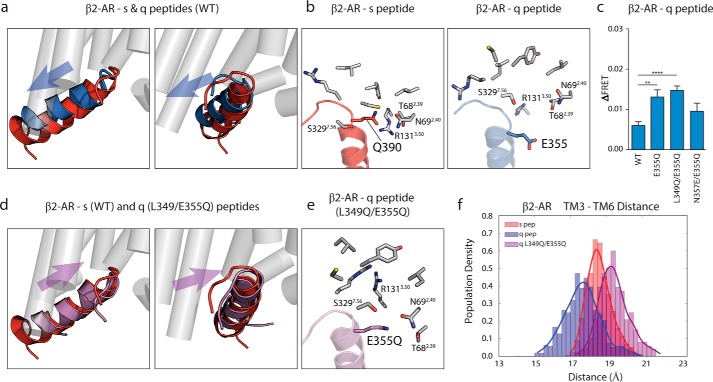
Although the importance of the C terminus of the α subunit of the heterotrimeric G protein in G protein-coupled receptor (GPCR)-G protein pairing is well established, the structural basis of selective interactions remains unknown. Here, we combine live cell FRET-based measurements and molecular dynamics simulations of the interaction between the GPCR and a peptide derived from the C terminus of the Gα subunit (Gα peptide) to dissect the molecular mechanisms of G protein selectivity. We observe a direct link between Gα peptide binding and stabilization of the GPCR conformational ensemble. We find that cognate and non-cognate Gα peptides show deep and shallow binding, respectively, and in distinct orientations within the GPCR. Binding of the cognate Gα peptide stabilizes the agonist-bound GPCR conformational ensemble resulting in favorable binding energy and lower flexibility of the agonist-GPCR pair. We identify three hot spot residues (Gαs/Gαq-Gln-384/Leu-349, Gln-390/Glu-355, and Glu-392/Asn-357) that contribute to selective interactions between the β2-adrenergic receptor (β2-AR)-Gαs and V1A receptor (V1AR)-Gαq The Gαs and Gαq peptides adopt different orientations in β2-AR and V1AR, respectively. The β2-AR/Gαs peptide interface is dominated by electrostatic interactions, whereas the V1AR/Gαq peptide interactions are predominantly hydrophobic. Interestingly, our study reveals a role for both favorable and unfavorable interactions in G protein selection. Residue Glu-355 in Gαq prevents this peptide from interacting strongly with β2-AR. Mutagenesis to the Gαs counterpart (E355Q) imparts a cognate-like interaction. Overall, our study highlights the synergy in molecular dynamics and FRET-based approaches to dissect the structural basis of selective G protein interactions.
Keywords: G protein; G protein-coupled receptor (GPCR); Receptor Conformation; cell signaling; fluorescence resonance energy transfer (FRET); molecular dynamics.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
Similar articles
-
Priming GPCR signaling through the synergistic effect of two G proteins.Proc Natl Acad Sci U S A. 2017 Apr 4;114(14):3756-3761. doi: 10.1073/pnas.1617232114. Epub 2017 Mar 21. Proc Natl Acad Sci U S A. 2017. PMID: 28325873 Free PMC article.
-
Minute-scale persistence of a GPCR conformation state triggered by non-cognate G protein interactions primes signaling.Nat Commun. 2019 Oct 23;10(1):4836. doi: 10.1038/s41467-019-12755-9. Nat Commun. 2019. PMID: 31645561 Free PMC article.
-
Conformational plasticity of the intracellular cavity of GPCR-G-protein complexes leads to G-protein promiscuity and selectivity.Proc Natl Acad Sci U S A. 2019 Jun 11;116(24):11956-11965. doi: 10.1073/pnas.1820944116. Epub 2019 May 28. Proc Natl Acad Sci U S A. 2019. PMID: 31138704 Free PMC article.
-
Strike a pose: Gαq complexes at the membrane.Trends Pharmacol Sci. 2014 Jan;35(1):23-30. doi: 10.1016/j.tips.2013.10.008. Epub 2013 Nov 26. Trends Pharmacol Sci. 2014. PMID: 24287282 Free PMC article. Review.
-
Decoding Gαq signaling.Life Sci. 2016 May 1;152:99-106. doi: 10.1016/j.lfs.2016.03.037. Epub 2016 Mar 22. Life Sci. 2016. PMID: 27012764 Review.
Cited by
-
Dynamic spatiotemporal determinants modulate GPCR:G protein coupling selectivity and promiscuity.Nat Commun. 2022 Dec 2;13(1):7428. doi: 10.1038/s41467-022-34055-5. Nat Commun. 2022. PMID: 36460632 Free PMC article.
-
Molecular Dynamics Simulations of the Allosteric Modulation of the Adenosine A2A Receptor by a Mini-G Protein.Sci Rep. 2019 Apr 2;9(1):5495. doi: 10.1038/s41598-019-41980-x. Sci Rep. 2019. PMID: 30940903 Free PMC article.
-
Priming GPCR signaling through the synergistic effect of two G proteins.Proc Natl Acad Sci U S A. 2017 Apr 4;114(14):3756-3761. doi: 10.1073/pnas.1617232114. Epub 2017 Mar 21. Proc Natl Acad Sci U S A. 2017. PMID: 28325873 Free PMC article.
-
Allosteric modulation of adenosine A1 and cannabinoid 1 receptor signaling by G-peptides.Pharmacol Res Perspect. 2020 Dec;8(6):e00673. doi: 10.1002/prp2.673. Pharmacol Res Perspect. 2020. PMID: 33124765 Free PMC article.
-
Molecular mechanism of muscarinic acetylcholine receptor M3 interaction with Gq.Commun Biol. 2024 Mar 23;7(1):362. doi: 10.1038/s42003-024-06056-1. Commun Biol. 2024. PMID: 38521872 Free PMC article.
References
-
- Kobilka B., and Schertler G. F. (2008) New G-protein-coupled receptor crystal structures: insights and limitations. Trends Pharmacol. Sci. 29, 79–83 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
