

Identification of Multiple Proteins Coupling Transcriptional Gene Silencing to Genome Stability in Arabidopsis thaliana
- PMID: 27253878
- PMCID: PMC4890748
- DOI: 10.1371/journal.pgen.1006092
Identification of Multiple Proteins Coupling Transcriptional Gene Silencing to Genome Stability in Arabidopsis thaliana
Abstract
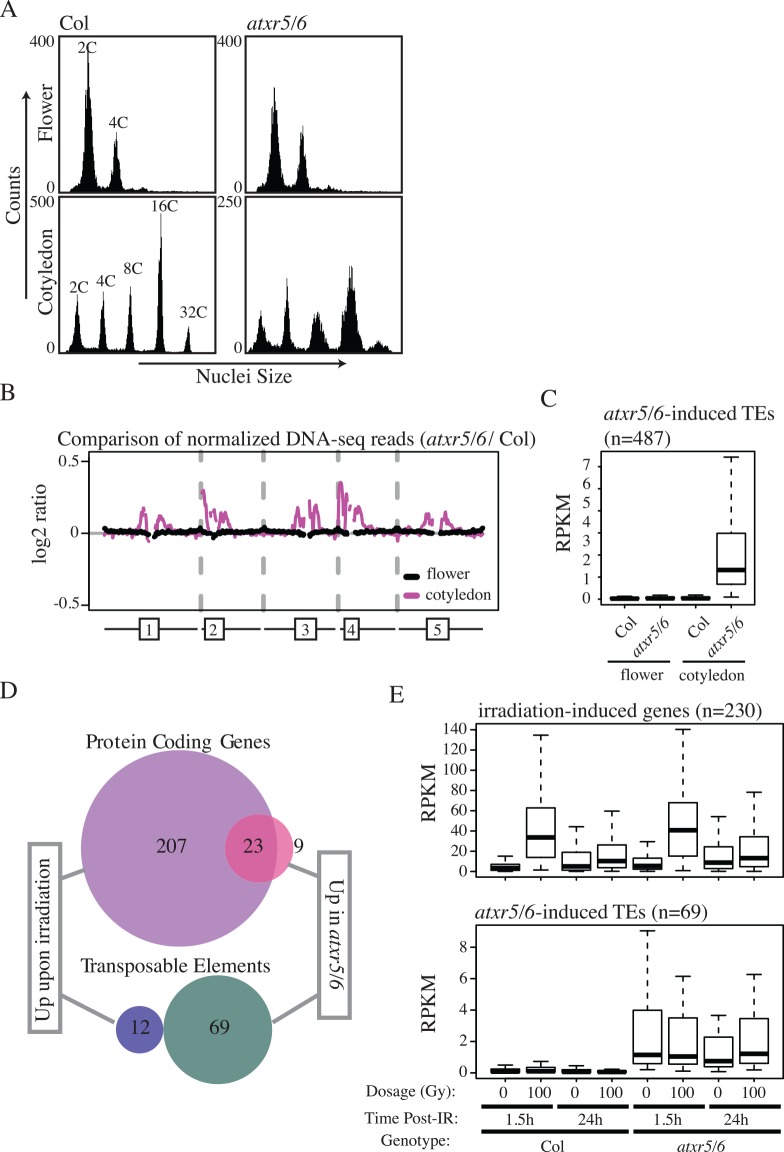
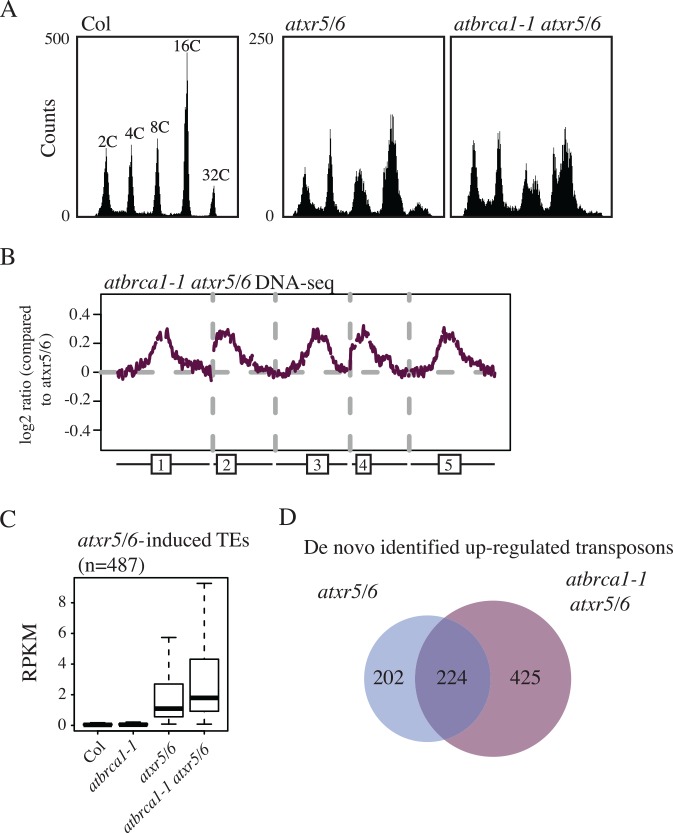
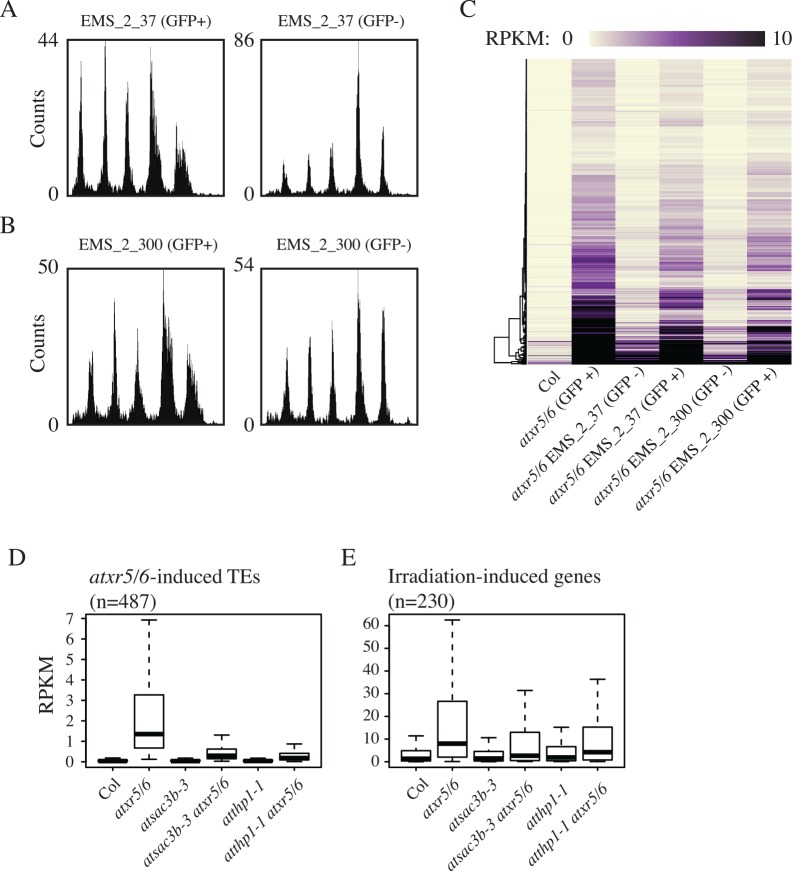
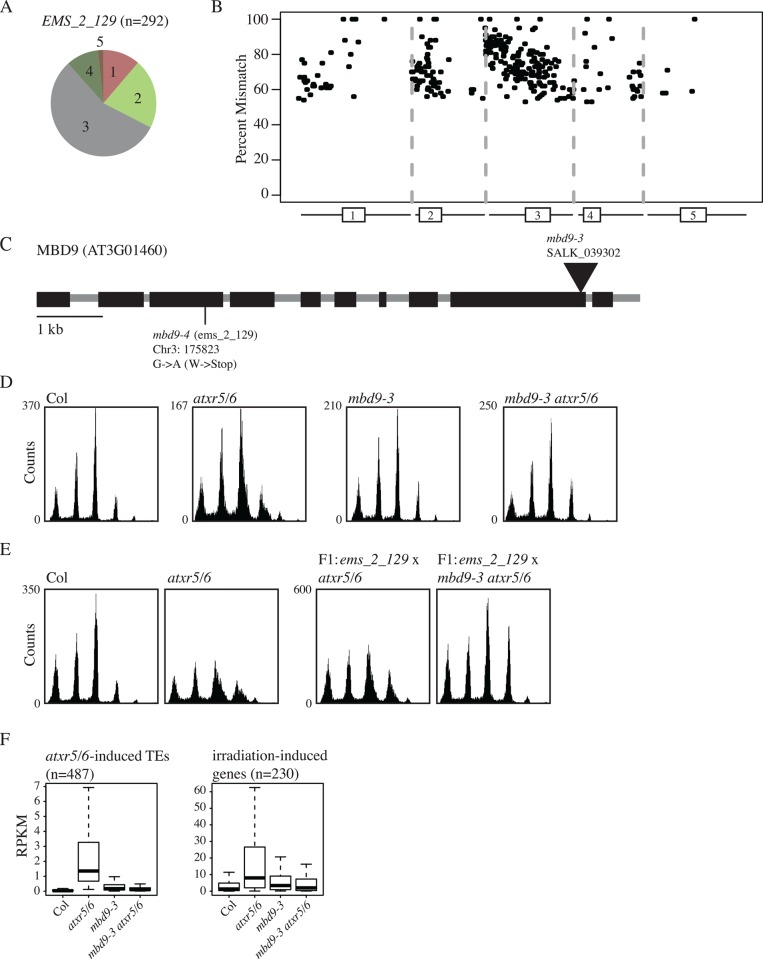
Eukaryotic genomes are regulated by epigenetic marks that act to modulate transcriptional control as well as to regulate DNA replication and repair. In Arabidopsis thaliana, mutation of the ATXR5 and ATXR6 histone methyltransferases causes reduction in histone H3 lysine 27 monomethylation, transcriptional upregulation of transposons, and a genome instability defect in which there is an accumulation of excess DNA corresponding to pericentromeric heterochromatin. We designed a forward genetic screen to identify suppressors of the atxr5/6 phenotype that uncovered loss-of-function mutations in two components of the TREX-2 complex (AtTHP1, AtSAC3B), a SUMO-interacting E3 ubiquitin ligase (AtSTUbL2) and a methyl-binding domain protein (AtMBD9). Additionally, using a reverse genetic approach, we show that a mutation in a plant homolog of the tumor suppressor gene BRCA1 enhances the atxr5/6 phenotype. Through characterization of these mutations, our results suggest models for the production atxr5 atxr6-induced extra DNA involving conflicts between the replicative and transcriptional processes in the cell, and suggest that the atxr5 atxr6 transcriptional defects may be the cause of the genome instability defects in the mutants. These findings highlight the critical intersection of transcriptional silencing and DNA replication in the maintenance of genome stability of heterochromatin.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
H3.1K27me1 maintains transcriptional silencing and genome stability by preventing GCN5-mediated histone acetylation.Plant Cell. 2021 May 31;33(4):961-979. doi: 10.1093/plcell/koaa027. Plant Cell. 2021. PMID: 33793815 Free PMC article.
-
Arabidopsis Serrate Coordinates Histone Methyltransferases ATXR5/6 and RNA Processing Factor RDR6 to Regulate Transposon Expression.Dev Cell. 2018 Jun 18;45(6):769-784.e6. doi: 10.1016/j.devcel.2018.05.023. Dev Cell. 2018. PMID: 29920280
-
Regulation of heterochromatic DNA replication by histone H3 lysine 27 methyltransferases.Nature. 2010 Aug 19;466(7309):987-91. doi: 10.1038/nature09290. Epub 2010 Jul 14. Nature. 2010. PMID: 20631708 Free PMC article.
-
Heterochromatin proteins and the control of heterochromatic gene silencing in Arabidopsis.J Plant Physiol. 2006 Feb;163(3):358-68. doi: 10.1016/j.jplph.2005.10.015. Epub 2005 Dec 27. J Plant Physiol. 2006. PMID: 16384625 Review.
-
Preventing transcriptional gene silencing by active DNA demethylation.FEBS Lett. 2005 Oct 31;579(26):5889-98. doi: 10.1016/j.febslet.2005.08.039. Epub 2005 Aug 31. FEBS Lett. 2005. PMID: 16162337 Review.
Cited by
-
Arabidopsis-expressing lysine-null SUMO1 reveals a non-essential role for secondary SUMO modifications in plants.Plant Direct. 2023 Jul 16;7(7):e506. doi: 10.1002/pld3.506. eCollection 2023 Jul. Plant Direct. 2023. PMID: 37465357 Free PMC article.
-
DNA polymerase epsilon is required for heterochromatin maintenance in Arabidopsis.Genome Biol. 2020 Nov 25;21(1):283. doi: 10.1186/s13059-020-02190-1. Genome Biol. 2020. PMID: 33234150 Free PMC article.
-
Identification of plants' functional counterpart of the metazoan mediator of DNA Damage checkpoint 1.EMBO Rep. 2024 Apr;25(4):1936-1961. doi: 10.1038/s44319-024-00107-8. Epub 2024 Mar 4. EMBO Rep. 2024. PMID: 38438802 Free PMC article.
-
Mutation of Arabidopsis SMC4 identifies condensin as a corepressor of pericentromeric transposons and conditionally expressed genes.Genes Dev. 2017 Aug 1;31(15):1601-1614. doi: 10.1101/gad.301499.117. Epub 2017 Sep 7. Genes Dev. 2017. PMID: 28882854 Free PMC article.
-
The making and unmaking of the silenced chromatin.Plant Cell. 2021 May 31;33(4):786-787. doi: 10.1093/plcell/koab001. Plant Cell. 2021. PMID: 35234961 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
