

Review
doi: 10.1186/s13059-016-0965-5.
Transposable elements in the mammalian embryo: pioneers surviving through stealth and service
Affiliations
- PMID: 27161170
- PMCID: PMC4862087
- DOI: 10.1186/s13059-016-0965-5
Item in Clipboard
Review
Transposable elements in the mammalian embryo: pioneers surviving through stealth and service
Genome Biol.
.
Abstract
Transposable elements (TEs) are notable drivers of genetic innovation. Over evolutionary time, TE insertions can supply new promoter, enhancer, and insulator elements to protein-coding genes and establish novel, species-specific gene regulatory networks. Conversely, ongoing TE-driven insertional mutagenesis, nonhomologous recombination, and other potentially deleterious processes can cause sporadic disease by disrupting genome integrity or inducing abrupt gene expression changes. Here, we discuss recent evidence suggesting that TEs may contribute regulatory innovation to mammalian embryonic and pluripotent states as a means to ward off complete repression by their host genome.
Figures
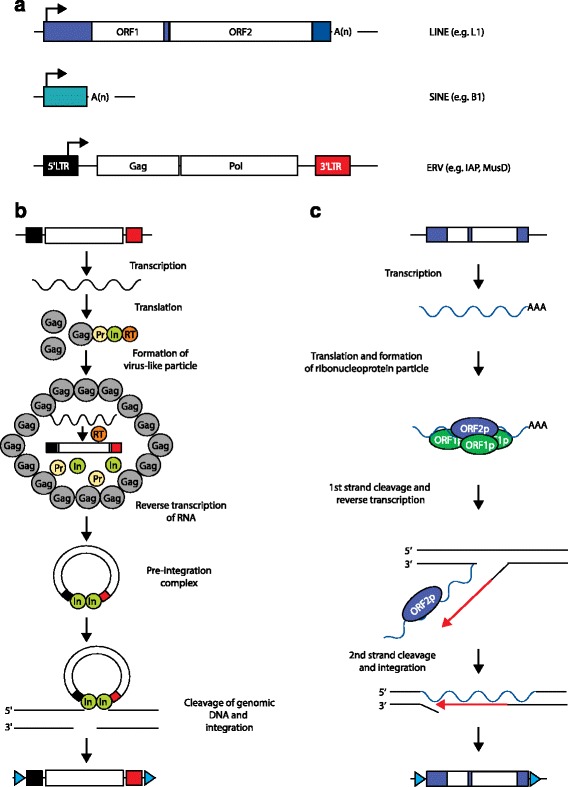
Long terminal repeat (LTR) and non-LTR retrotransposition mechanisms. a Mammalian retrotransposon structures. A long interspersed element (LINE; human L1 shown) typically consists of a 5′ untranslated region (UTR; blue box) harboring an internal promoter, two open reading frames (ORF1, ORF2), a 3′ UTR (small blue box), and a poly(A)-tail. A short interspersed element (SINE; mouse B1 shown) does not encode proteins and is trans-mobilized by LINE proteins. An endogenous retrovirus (ERV), such as mouse intracisternal A-type particle (IAP) and Mus type-D related retrovirus (MusD), lacks an Env protein but encodes functional Gag and Pol proteins flanked by a LTR at the 5′ (black box) and 3′ (red box) ends. Arrows indicate transcription start sites. b ERV mobilization starts with mRNA transcription and translation to yield Gag and Gag–Pro–Pol fusion proteins. The fusion proteins consist of a Gag protein (Gag), a protease (Pr), an integrase (In), and a reverse transcriptase (RT). Gag proteins build a virus-like particle and encapsulate the fusion proteins, which are processed into separate mature proteins. The ERV mRNA is then reverse transcribed, generating a cDNA. This cDNA and the integrase build a preintegration complex. The integrase then creates a double-strand DNA break, followed by genomic integration of a new ERV copy. Target site duplications (TSDs) are indicated by blue triangles. c L1 mobilization begins with transcription of an L1 mRNA, which is translated to yield ORF1p and ORF2p. ORF1p, ORF2p, and the L1 mRNA form a ribonucleoprotein particle that re-enters the nucleus. The ORF2p endonuclease cleaves the first genomic DNA strand, while its reverse transcriptase uses a now free 3′ OH group as a primer for reverse transcription of the L1 mRNA. Following second-strand DNA cleavage, a new L1 copy is integrated into the genome and is typically flanked by TSDs
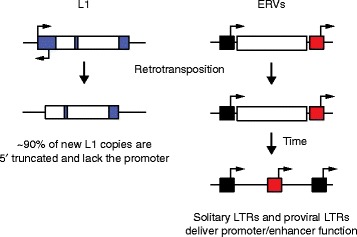
Long interspersed element 1 (L1) and endogenous retrovirus (ERV) regulatory impact post-integration. Most L1 copies are 5′ truncated (left) and lack the sense and antisense L1 promoters located in the 5′ untranslated region (large blue box). As a result, these L1 insertions have less capacity to drive chimeric transcription with neighboring genes. ERV insertions (right) remain either full-length, with flanking 5′ (black box) and 3′ long terminal repeats (LTRs; red box) that potentially retain promoter function, or, more commonly, recombine between the LTRs to form a solitary LTR, which retains the promoter/enhancer region. Arrows indicate putative transcription start sites
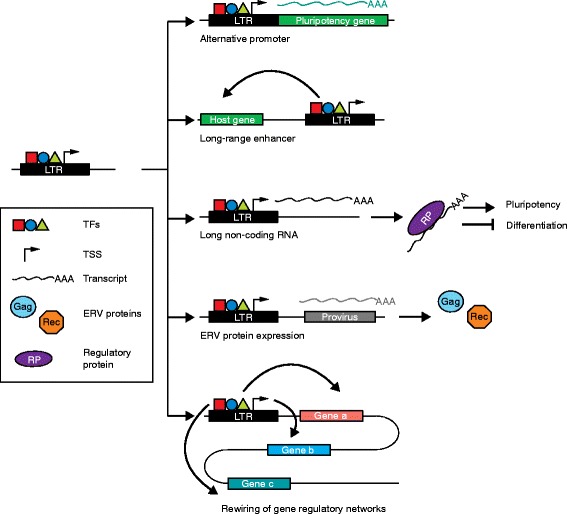
Examples of endogenous retrovirus (ERV) contributions to pluripotency. A long terminal repeat (LTR) possesses binding sites for pluripotency transcription factors (TFs) and can serve as a transcription start site (TSS). LTRs bound by pluripotency TFs can thereby impact embryonic stem cell identity by: (1) serving as alternative promoters for pluripotency genes, (2) providing long-range enhancers to specific host genes, (3) generating stem-cell-specific long noncoding RNAs that can bind to proteins regulating the pluripotent state, (4) transcribing proviral DNA elements as precursors to ERV protein expression, and (5) rewiring gene regulatory networks by controlling several pluripotency genes
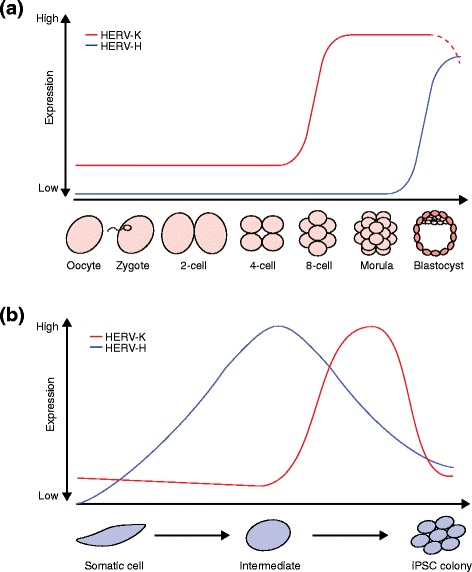
Human endogenous retrovirus (HERV) expression patterns in pluripotent cells. a HERV-K transcription in human embryogenesis is initiated during embryonic genome activation at the eight-cell stage and remains until the blastocyst stage. Dashed lines indicate proposed expression of HERV-K [31]. HERV-H can only be detected in epiblast cells of the late blastocyst [25]. b After induction of induced pluripotent stem cell (iPSC) reprogramming, HERV-K and HERV-H are derepressed with distinct dynamics. HERV-K transcription reaches its peak shortly before cells are fully reprogrammed. HERV-K expression subsequently decreases in reprogrammed cells and is silenced in iPSCs [32]. HERV-H is highly expressed earlier during reprogramming compared with HERV-K [24]. Note: the time points shown are approximate due to technical differences between studies
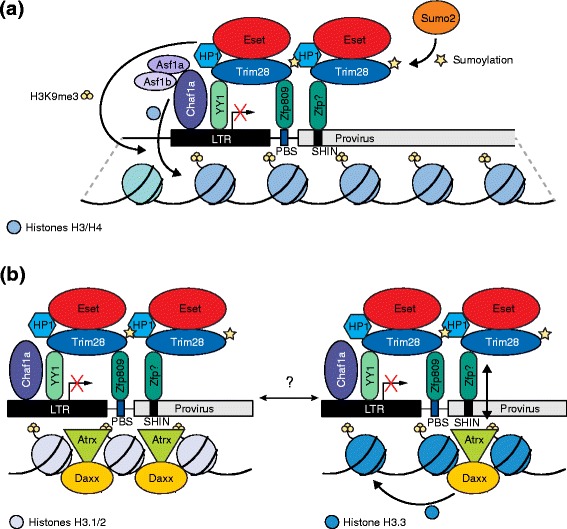
Proposed models of de novo endogenous retrovirus (ERV) silencing in embryonic stem cells. a To initiate silencing, the Krüppel-associated box (KRAB) zinc finger protein (Zfp) Zfp809 interacts with the proline primer binding site (PBS Pro) of some ERV families (e.g., Moloney murine leukemia virus) [85] whereas other KRAB-Zfps bind to a short heterochromatin-inducing (SHIN) sequence found in intracisternal A-type particle retrotransposons and other ERV families [93]. Subsequently, Trim28 is recruited by the Zfps [74, 86], assisted by binding of YY1 to the long terminal repeat (LTR) and Trim28 [92]. Interaction with HP1 and sumolyation by Sumo2 are thought to contribute to transcriptional repression mediated by Trim28 [72, 86, 89]. Eset also interacts with Trim28 and enables trimethylation of H3K9 and H4K20 [73]. The histone chaperone Chaf1a, aided by Asf1a/b, marks proviral DNA for silencing by depositing histones H3 and H4 and interacts with Eset [72]. b Conflicting models of ERV silencing by H3.3 deposition. The Atrx–Daxx complex is suggested to play an important role in SHIN-mediated silencing, which is H3.3-independent. Here, Atrx is thought to promote ERV heterochromatin inaccessibility (left) [93]. However, Atrx–Daxx is also proposed to deposit H3.3 and to interact with Trim28, followed by H3.3 being marked with H3K9me3 by Eset (right) [95]
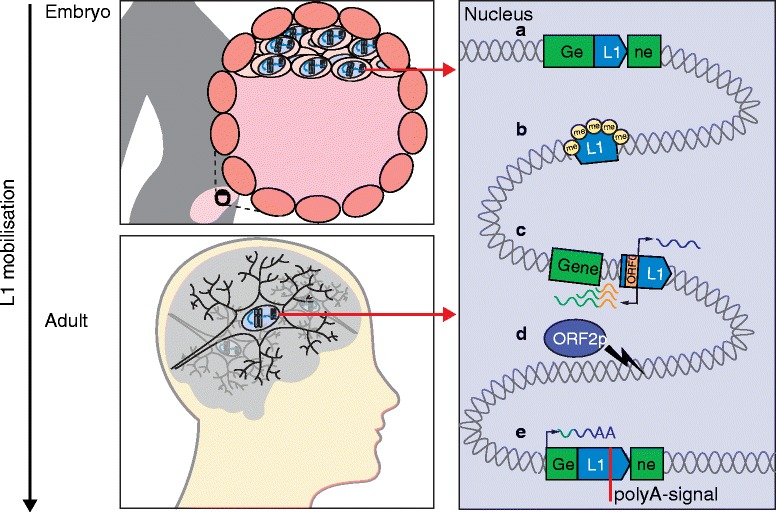
Long interspersed element-1 (L1) contributes to somatic mosaicism. L1 mobilizes in the brain and early embryo (left) and may, for example: a insert into protein-coding exons; b influence neighboring genes by the spreading of repressive histone modifications, such as methylation (me); c initiate sense or antisense transcription of neighboring genes, thereby creating new transcripts, including open reading frame 0 (ORF0) fusion transcripts, using host gene provided splice acceptor sites, which are translated to fusion proteins; d generate DNA double-strand breaks via the endonuclease activity of L1 ORF2p; and e lead to premature termination of host gene transcripts by providing alternative poly(A) signals
Similar articles
-
The developmental control of transposable elements and the evolution of higher species.Annu Rev Cell Dev Biol. 2015;31:429-51. doi: 10.1146/annurev-cellbio-100814-125514. Epub 2015 Sep 17. Annu Rev Cell Dev Biol. 2015. PMID: 26393776 Review.
-
Functional evaluation of transposable elements as enhancers in mouse embryonic and trophoblast stem cells.Elife. 2019 Apr 23;8:e44344. doi: 10.7554/eLife.44344. Elife. 2019. PMID: 31012843 Free PMC article.
-
Transposable elements as genetic regulatory substrates in early development.Trends Cell Biol. 2013 May;23(5):218-26. doi: 10.1016/j.tcb.2013.01.001. Epub 2013 Feb 12. Trends Cell Biol. 2013. PMID: 23411159 Free PMC article. Review.
-
Host Gene Regulation by Transposable Elements: The New, the Old and the Ugly.Viruses. 2020 Sep 26;12(10):1089. doi: 10.3390/v12101089. Viruses. 2020. PMID: 32993145 Free PMC article. Review.
-
Technology to the rescue: how to uncover the role of transposable elements in preimplantation development.Biochem Soc Trans. 2024 Jun 26;52(3):1349-1362. doi: 10.1042/BST20231262. Biochem Soc Trans. 2024. PMID: 38752836 Free PMC article. Review.
Cited by
-
Trim24 and Trim33 Play a Role in Epigenetic Silencing of Retroviruses in Embryonic Stem Cells.Viruses. 2020 Sep 11;12(9):1015. doi: 10.3390/v12091015. Viruses. 2020. PMID: 32932986 Free PMC article.
-
Transcript assembly improves expression quantification of transposable elements in single-cell RNA-seq data.Genome Res. 2021 Jan;31(1):88-100. doi: 10.1101/gr.265173.120. Epub 2020 Dec 21. Genome Res. 2021. PMID: 33355230 Free PMC article.
-
The coevolution between APOBEC3 and retrotransposons in primates.Mob DNA. 2022 Nov 29;13(1):27. doi: 10.1186/s13100-022-00283-1. Mob DNA. 2022. PMID: 36443831 Free PMC article. Review.
-
CTRL+INSERT: retrotransposons and their contribution to regulation and innovation of the transcriptome.EMBO Rep. 2016 Aug;17(8):1131-44. doi: 10.15252/embr.201642743. Epub 2016 Jul 11. EMBO Rep. 2016. PMID: 27402545 Free PMC article. Review.
-
Reactivation of a somatic errantivirus and germline invasion in Drosophila ovaries.Nat Commun. 2023 Sep 29;14(1):6096. doi: 10.1038/s41467-023-41733-5. Nat Commun. 2023. PMID: 37773253 Free PMC article.
References
-
- Papaioannou VE, Mkandawire J, Biggers JD. Development and phenotypic variability of genetically identical half mouse embryos. Development. 1989;106:817–27. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
