

Biofilms and Cyclic di-GMP (c-di-GMP) Signaling: Lessons from Pseudomonas aeruginosa and Other Bacteria
- PMID: 27129226
- PMCID: PMC4933438
- DOI: 10.1074/jbc.R115.711507
Biofilms and Cyclic di-GMP (c-di-GMP) Signaling: Lessons from Pseudomonas aeruginosa and Other Bacteria
Abstract
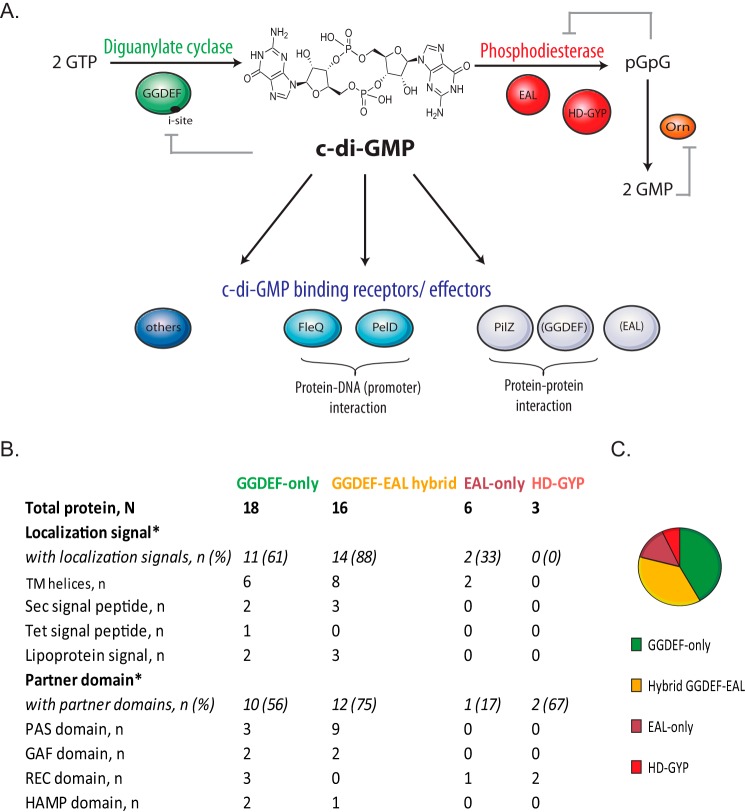
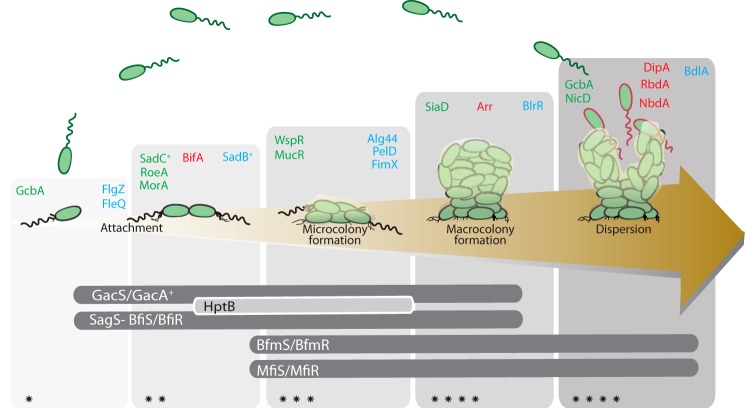
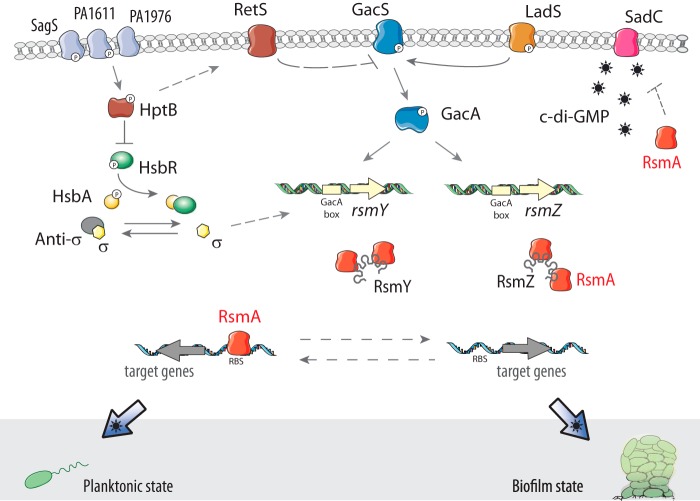
The cyclic di-GMP (c-di-GMP) second messenger represents a signaling system that regulates many bacterial behaviors and is of key importance for driving the lifestyle switch between motile loner cells and biofilm formers. This review provides an up-to-date compendium of c-di-GMP pathways connected to biofilm formation, biofilm-associated motilities, and other functionalities in the ubiquitous and opportunistic human pathogen Pseudomonas aeruginosa This bacterium is frequently adopted as a model organism to study bacterial biofilm formation. Importantly, its versatility and adaptation capabilities are linked with a broad range of complex regulatory networks, including a large set of genes involved in c-di-GMP biosynthesis, degradation, and transmission.
Keywords: Pseudomonas aeruginosa (P. aeruginosa); antibiotic resistance; biofilm; cyclic di-GMP (c-di-GMP); signaling.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
Similar articles
-
Controlling Biofilm Development Through Cyclic di-GMP Signaling.Adv Exp Med Biol. 2022;1386:69-94. doi: 10.1007/978-3-031-08491-1_3. Adv Exp Med Biol. 2022. PMID: 36258069 Free PMC article. Review.
-
Heterogeneity in surface sensing suggests a division of labor in Pseudomonas aeruginosa populations.Elife. 2019 Jun 10;8:e45084. doi: 10.7554/eLife.45084. Elife. 2019. PMID: 31180327 Free PMC article.
-
Glucose-6-Phosphate Acts as an Extracellular Signal of SagS To Modulate Pseudomonas aeruginosa c-di-GMP Levels, Attachment, and Biofilm Formation.mSphere. 2021 Feb 10;6(1):e01231-20. doi: 10.1128/mSphere.01231-20. mSphere. 2021. PMID: 33568456 Free PMC article.
-
A Cyclic di-GMP-binding Adaptor Protein Interacts with Histidine Kinase to Regulate Two-component Signaling.J Biol Chem. 2016 Jul 29;291(31):16112-23. doi: 10.1074/jbc.M116.730887. Epub 2016 May 26. J Biol Chem. 2016. PMID: 27231351 Free PMC article.
-
A PilZ domain protein for chemotaxis adds another layer to c-di-GMP-mediated regulation of flagellar motility.Sci Signal. 2016 Oct 18;9(450):fs16. doi: 10.1126/scisignal.aai8859. Sci Signal. 2016. PMID: 27811181
Cited by
-
The Role of Efflux and Physiological Adaptation in Biofilm Tolerance and Resistance.J Biol Chem. 2016 Jun 10;291(24):12565-12572. doi: 10.1074/jbc.R115.707257. Epub 2016 Apr 21. J Biol Chem. 2016. PMID: 27129224 Free PMC article. Review.
-
What's on the Outside Matters: The Role of the Extracellular Polymeric Substance of Gram-negative Biofilms in Evading Host Immunity and as a Target for Therapeutic Intervention.J Biol Chem. 2016 Jun 10;291(24):12538-12546. doi: 10.1074/jbc.R115.707547. Epub 2016 Apr 21. J Biol Chem. 2016. PMID: 27129225 Free PMC article. Review.
-
Microbial Biofilms in Pulmonary and Critical Care Diseases.Ann Am Thorac Soc. 2016 Sep;13(9):1615-23. doi: 10.1513/AnnalsATS.201603-194FR. Ann Am Thorac Soc. 2016. PMID: 27348071 Free PMC article. Review.
-
Degradation of cyclic diguanosine monophosphate by a hybrid two-component protein protects Azoarcus sp. strain CIB from toluene toxicity.Proc Natl Acad Sci U S A. 2016 Nov 15;113(46):13174-13179. doi: 10.1073/pnas.1615981113. Epub 2016 Oct 31. Proc Natl Acad Sci U S A. 2016. PMID: 27799551 Free PMC article.
-
Understanding Biofilms and Novel Approaches to the Diagnosis, Prevention, and Treatment of Medical Device-Associated Infections.Infect Dis Clin North Am. 2018 Dec;32(4):915-929. doi: 10.1016/j.idc.2018.06.009. Epub 2018 Sep 18. Infect Dis Clin North Am. 2018. PMID: 30241715 Free PMC article. Review.
References
-
- O'Toole G., Kaplan H. B., and Kolter R. (2000) Biofilm formation as microbial development. Annu. Rev. Microbiol. 54, 49–79 - PubMed
-
- Hall-Stoodley L., Costerton J. W., and Stoodley P. (2004) Bacterial biofilms: from the natural environment to infectious diseases. Nat. Rev. Microbiol. 2, 95–108 - PubMed
-
- Simm R., Morr M., Kader A., Nimtz M., and Römling U. (2004) GGDEF and EAL domains inversely regulate cyclic di-GMP levels and transition from sessility to motility. Mol. Microbiol. 53, 1123–1134 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
