

Repulsive cues combined with physical barriers and cell-cell adhesion determine progenitor cell positioning during organogenesis
- PMID: 27088892
- PMCID: PMC4837475
- DOI: 10.1038/ncomms11288
Repulsive cues combined with physical barriers and cell-cell adhesion determine progenitor cell positioning during organogenesis
Abstract
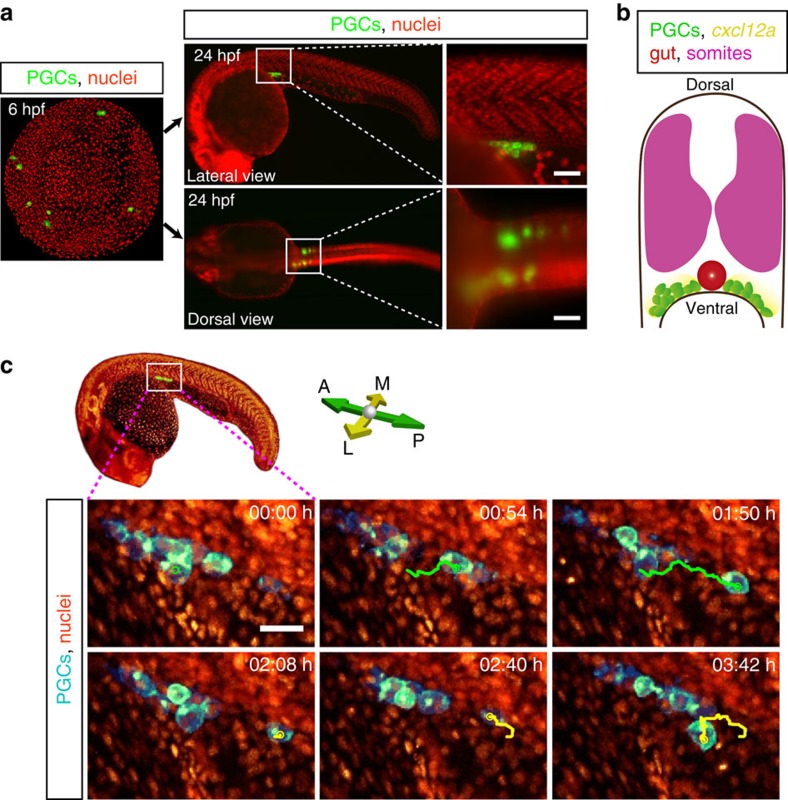
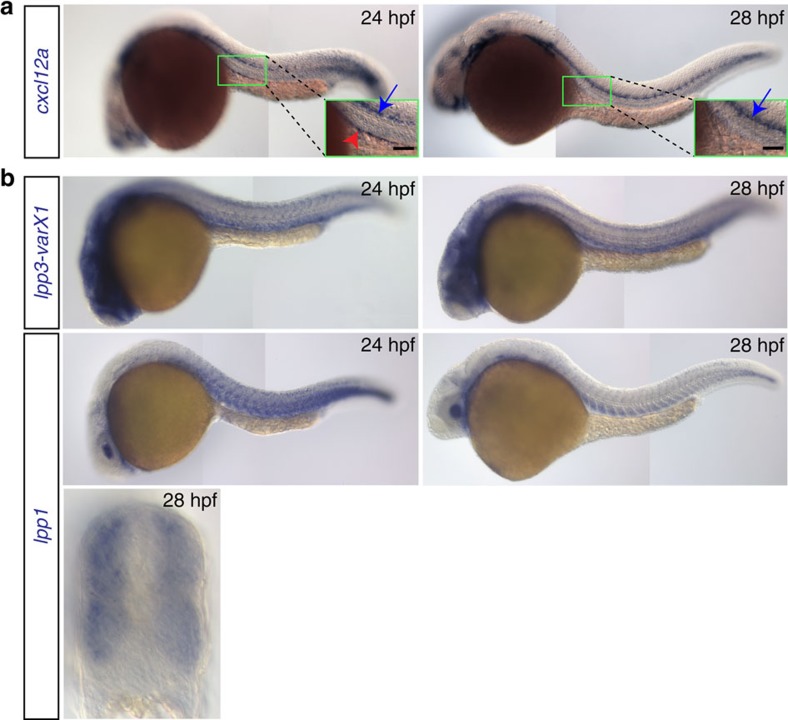
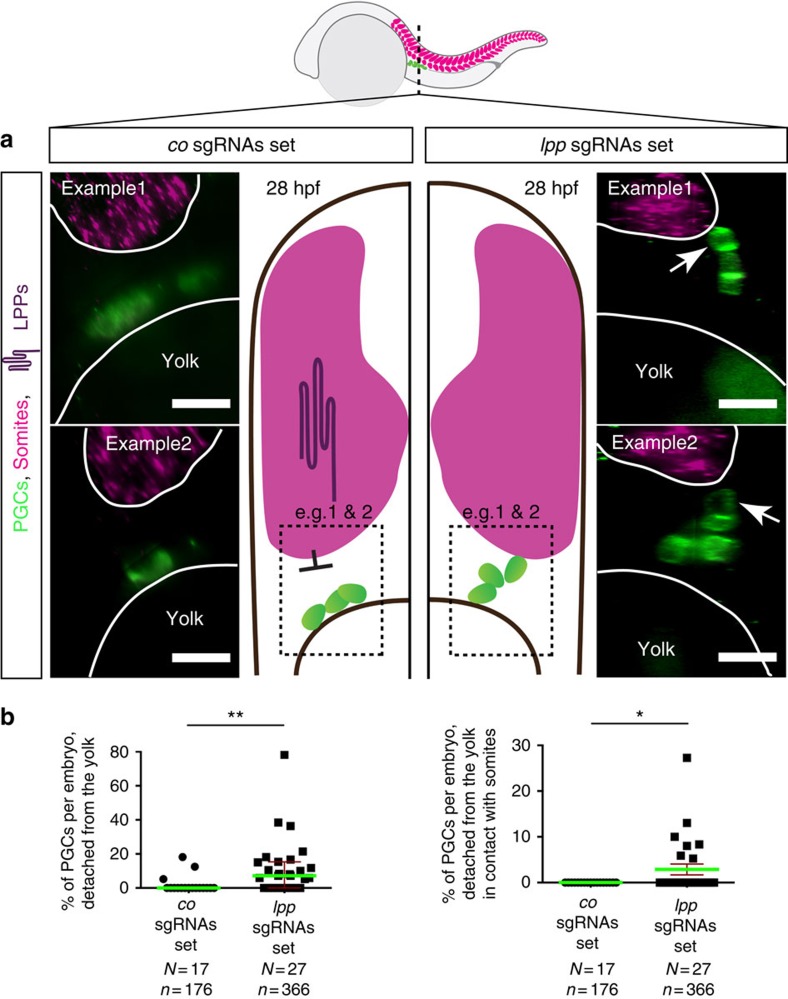
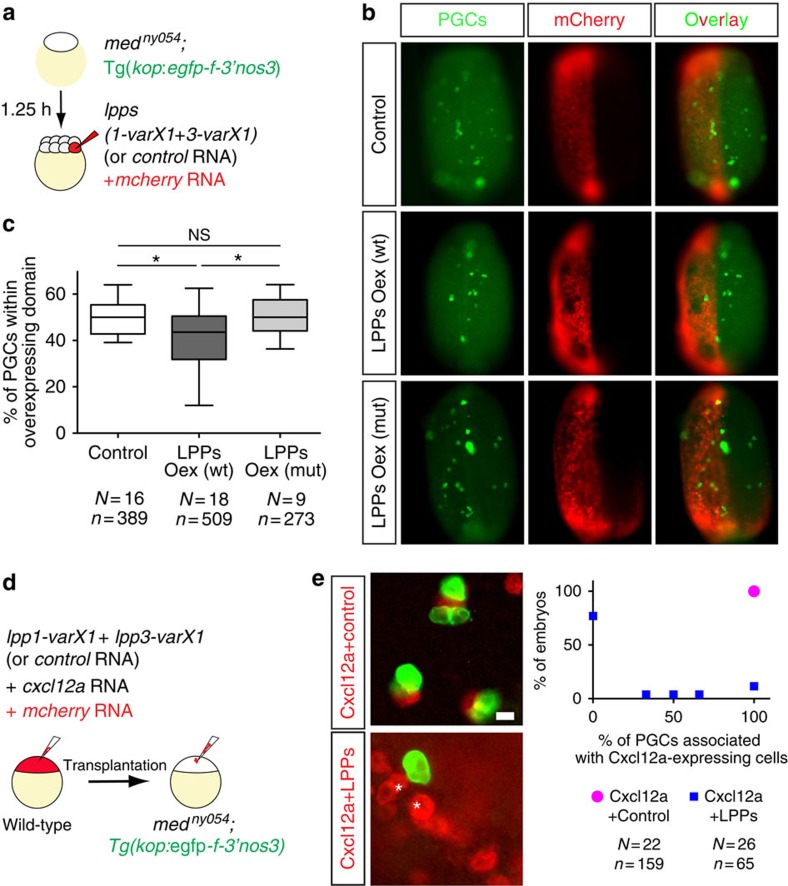
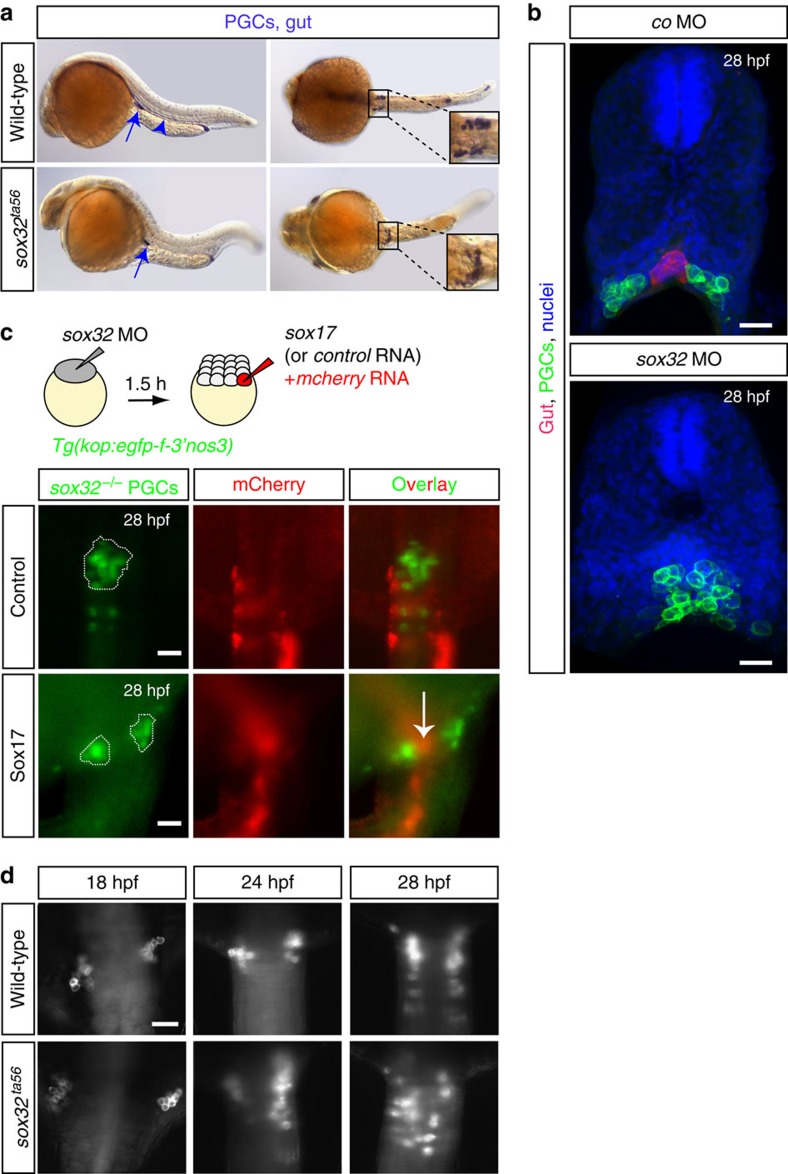
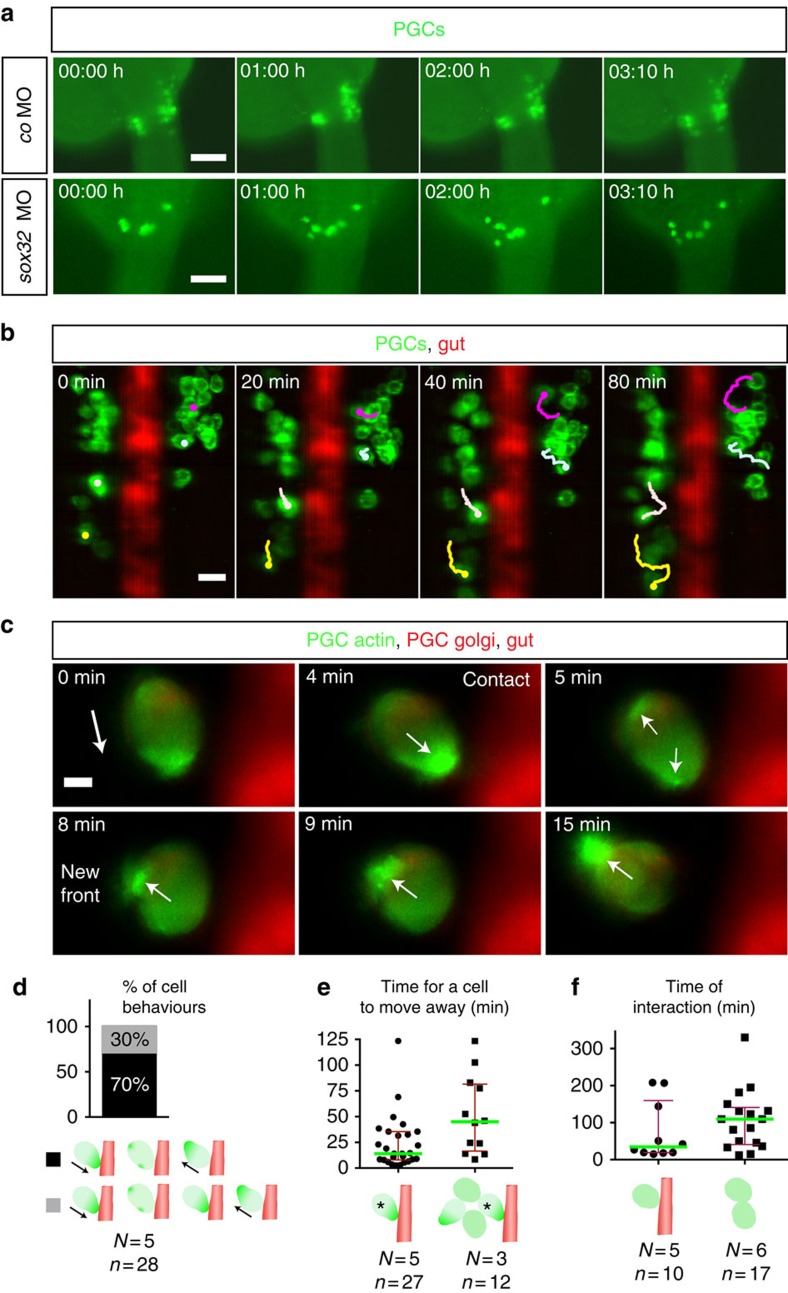
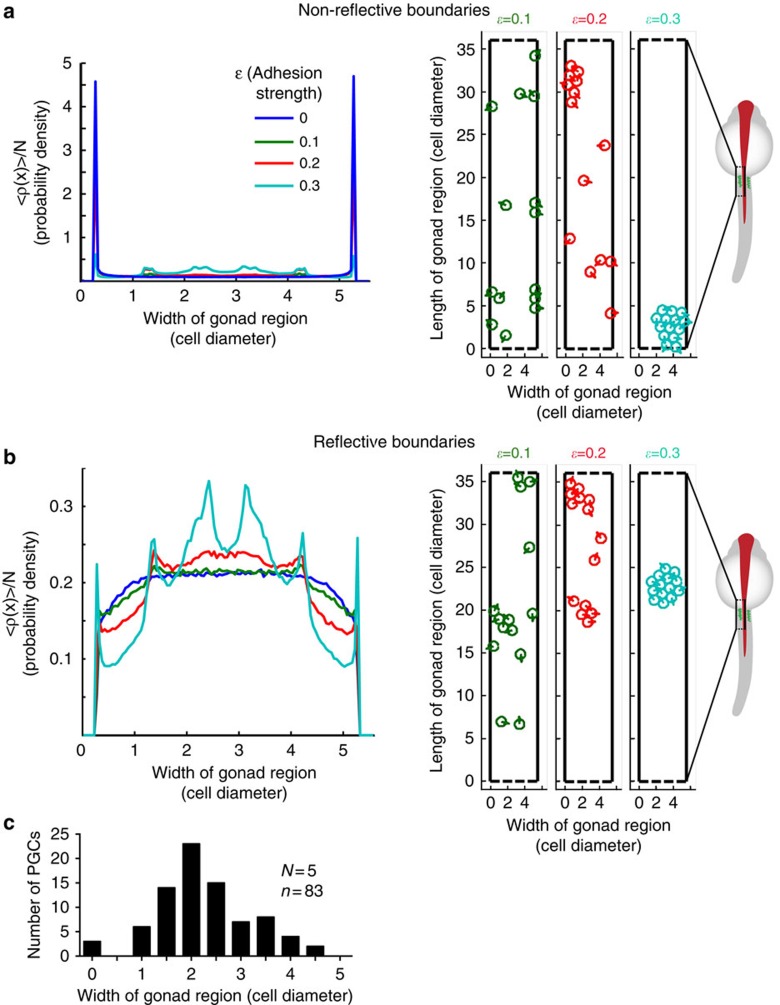
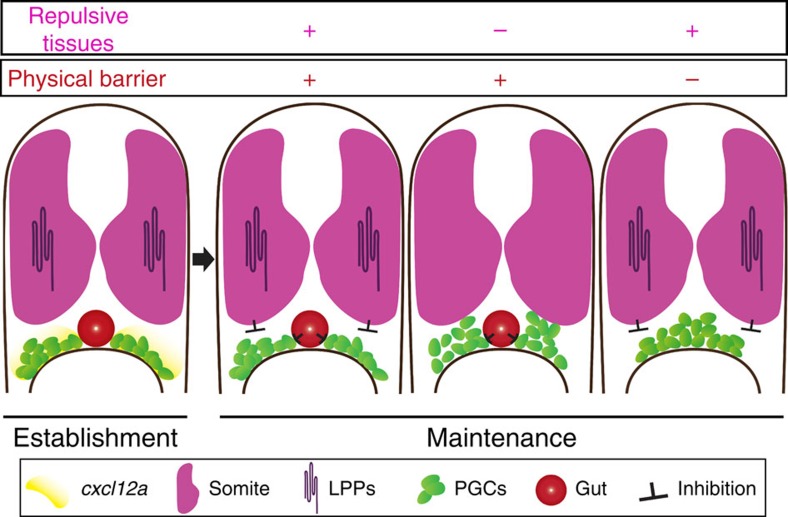
The precise positioning of organ progenitor cells constitutes an essential, yet poorly understood step during organogenesis. Using primordial germ cells that participate in gonad formation, we present the developmental mechanisms maintaining a motile progenitor cell population at the site where the organ develops. Employing high-resolution live-cell microscopy, we find that repulsive cues coupled with physical barriers confine the cells to the correct bilateral positions. This analysis revealed that cell polarity changes on interaction with the physical barrier and that the establishment of compact clusters involves increased cell-cell interaction time. Using particle-based simulations, we demonstrate the role of reflecting barriers, from which cells turn away on contact, and the importance of proper cell-cell adhesion level for maintaining the tight cell clusters and their correct positioning at the target region. The combination of these developmental and cellular mechanisms prevents organ fusion, controls organ positioning and is thus critical for its proper function.
Figures
Similar articles
-
Morpholino knockdown of the ubiquitously expressed transmembrane serine protease TMPRSS4a in zebrafish embryos exhibits severe defects in organogenesis and cell adhesion.Biol Chem. 2011 Jul;392(7):653-64. doi: 10.1515/BC.2011.070. Biol Chem. 2011. PMID: 21657981
-
Spatiotemporal expression of smooth muscle markers in developing zebrafish gut.Dev Dyn. 2007 Jun;236(6):1623-32. doi: 10.1002/dvdy.21165. Dev Dyn. 2007. PMID: 17474123
-
Role of a novel zebrafish nup98 during embryonic development.Exp Hematol. 2010 Nov;38(11):1014-1021.e1-2. doi: 10.1016/j.exphem.2010.07.010. Epub 2010 Aug 7. Exp Hematol. 2010. PMID: 20696206
-
Fly fishing for collective cell migration.Curr Opin Genet Dev. 2010 Aug;20(4):428-32. doi: 10.1016/j.gde.2010.04.006. Epub 2010 May 6. Curr Opin Genet Dev. 2010. PMID: 20452199 Review.
-
Guidance of primordial germ cell migration.Curr Opin Cell Biol. 2004 Apr;16(2):169-73. doi: 10.1016/j.ceb.2004.01.004. Curr Opin Cell Biol. 2004. PMID: 15196560 Review.
Cited by
-
The journey of a generation: advances and promises in the study of primordial germ cell migration.Development. 2024 Apr 1;151(7):dev201102. doi: 10.1242/dev.201102. Epub 2024 Apr 12. Development. 2024. PMID: 38607588 Free PMC article.
-
Bridging from single to collective cell migration: A review of models and links to experiments.PLoS Comput Biol. 2020 Dec 10;16(12):e1008411. doi: 10.1371/journal.pcbi.1008411. eCollection 2020 Dec. PLoS Comput Biol. 2020. PMID: 33301528 Free PMC article. Review.
-
Cell size: a key determinant of meristematic potential in plant protoplasts.aBIOTECH. 2021 Jan 1;2(1):96-104. doi: 10.1007/s42994-020-00033-y. eCollection 2021 Mar. aBIOTECH. 2021. PMID: 36304480 Free PMC article. Review.
-
Fluid dynamics during bleb formation in migrating cells in vivo.PLoS One. 2019 Feb 26;14(2):e0212699. doi: 10.1371/journal.pone.0212699. eCollection 2019. PLoS One. 2019. PMID: 30807602 Free PMC article.
-
Using migrating cells as probes to illuminate features in live embryonic tissues.Sci Adv. 2020 Dec 4;6(49):eabc5546. doi: 10.1126/sciadv.abc5546. Print 2020 Dec. Sci Adv. 2020. PMID: 33277250 Free PMC article.
References
-
- Koopman P. Organogenesis in Development. Curr. Top. Dev. Biol. 90, 1–408 (2010). - PubMed
-
- Liu W., Wu R. & Guo Z. The diagnosis and management of continuous splenogonadal fusion in a 6-year-old boy. Int. Urol. Nephrol. 45, 21–24 (2012). - PubMed
-
- Ferro F. et al. Hepatogonadal fusion. J. Pediatr. Surg. 31, 435–436 (1996). - PubMed
-
- Ueno T. et al. Spectrum of germ cell tumors: from head to toe. Radiographics 387–404 (2004). - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
