

Defining functional classes of Barth syndrome mutation in humans
- PMID: 26908608
- PMCID: PMC4986330
- DOI: 10.1093/hmg/ddw046
Defining functional classes of Barth syndrome mutation in humans
Abstract
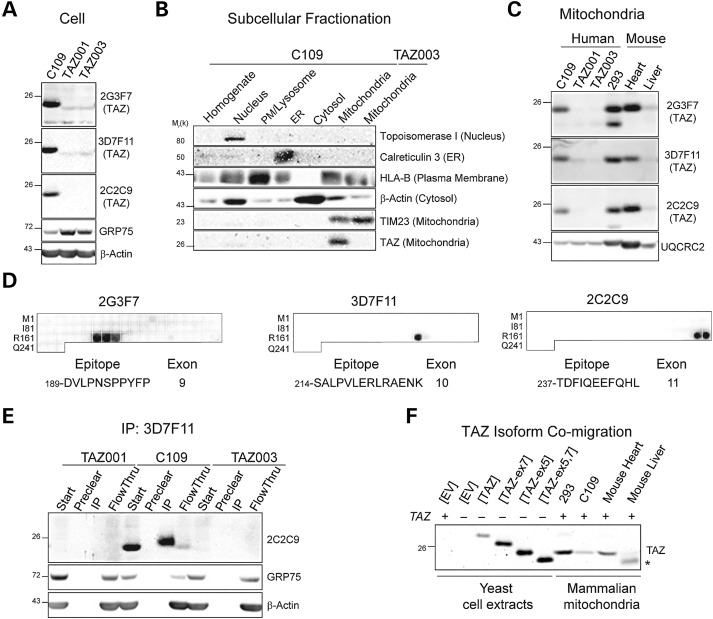
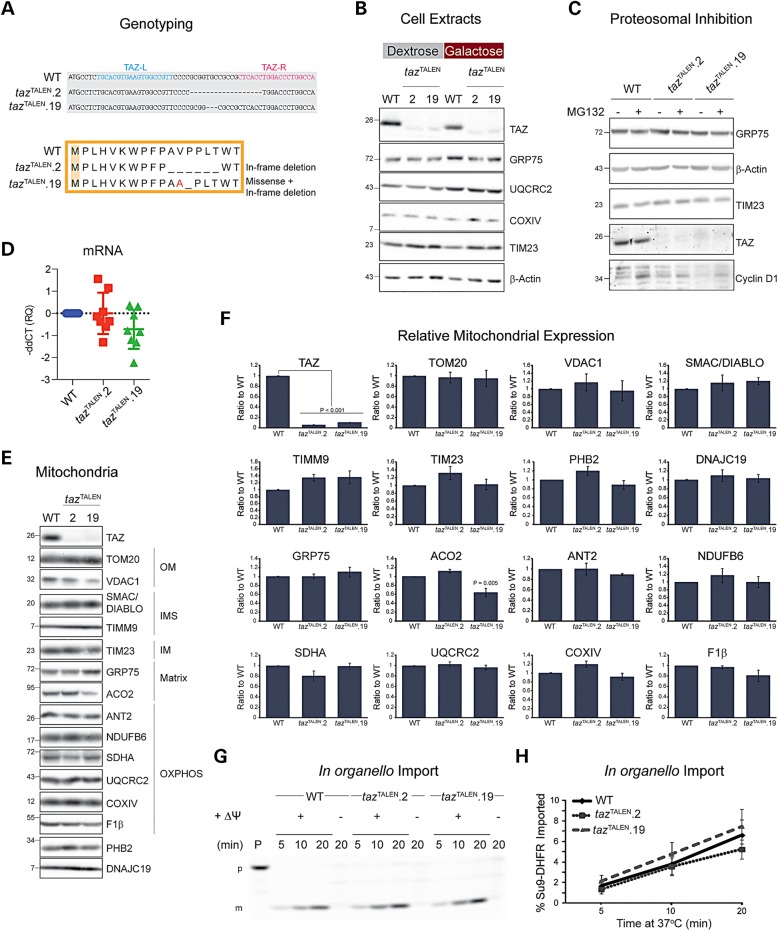
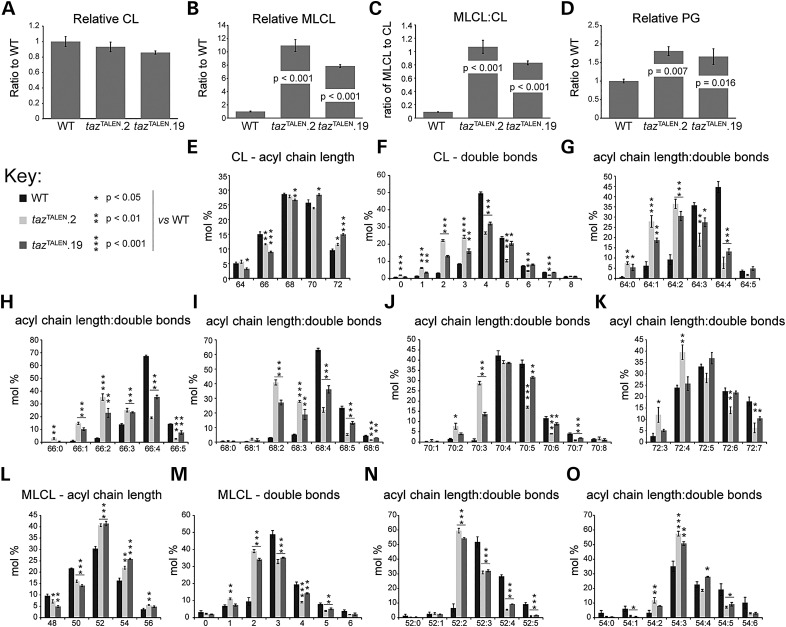
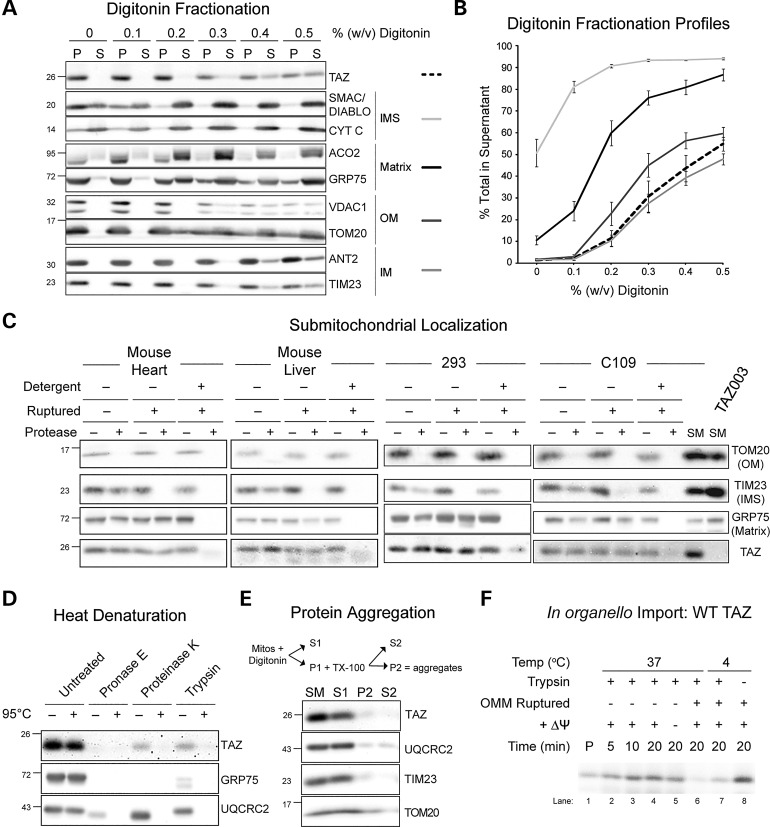
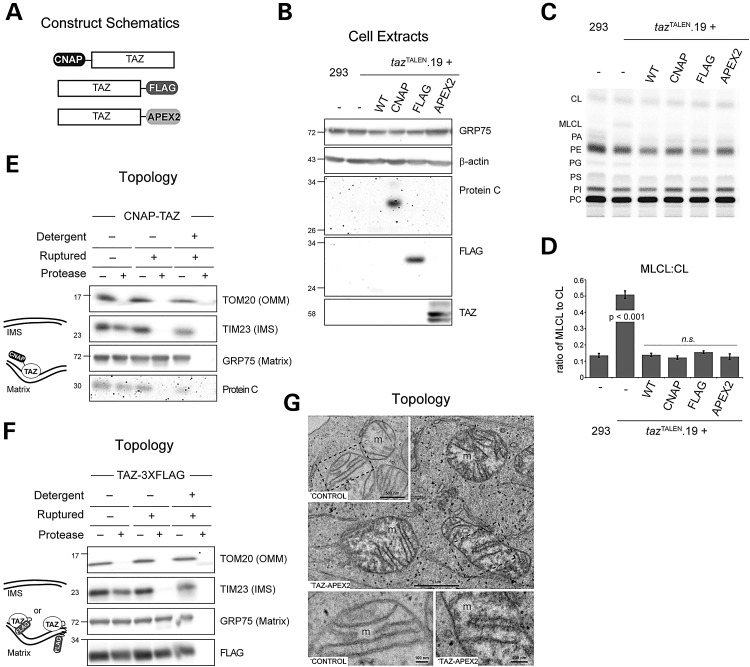
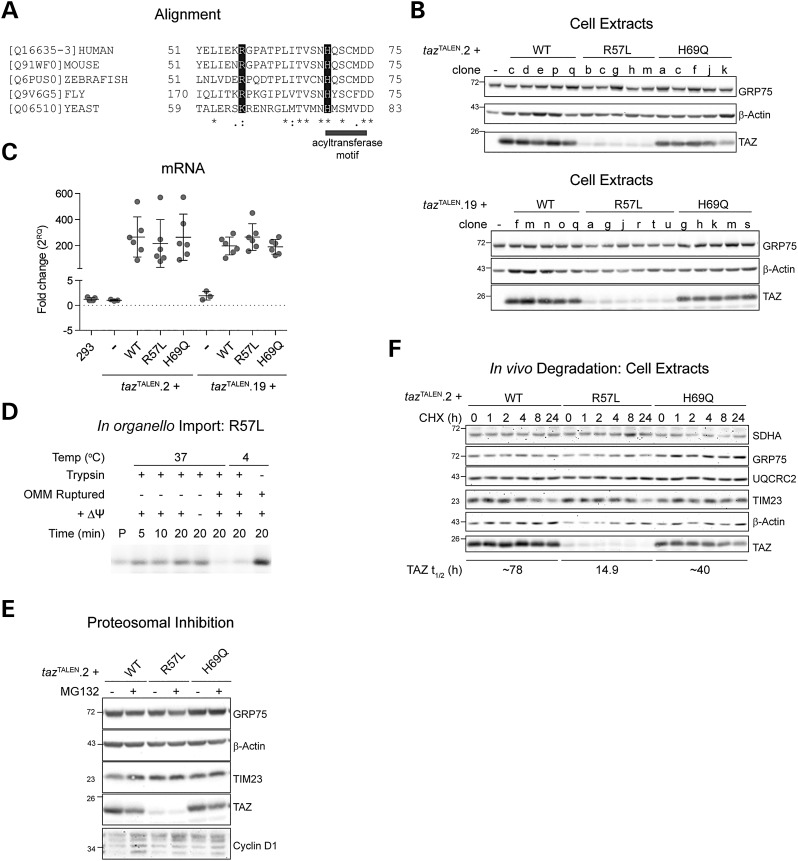
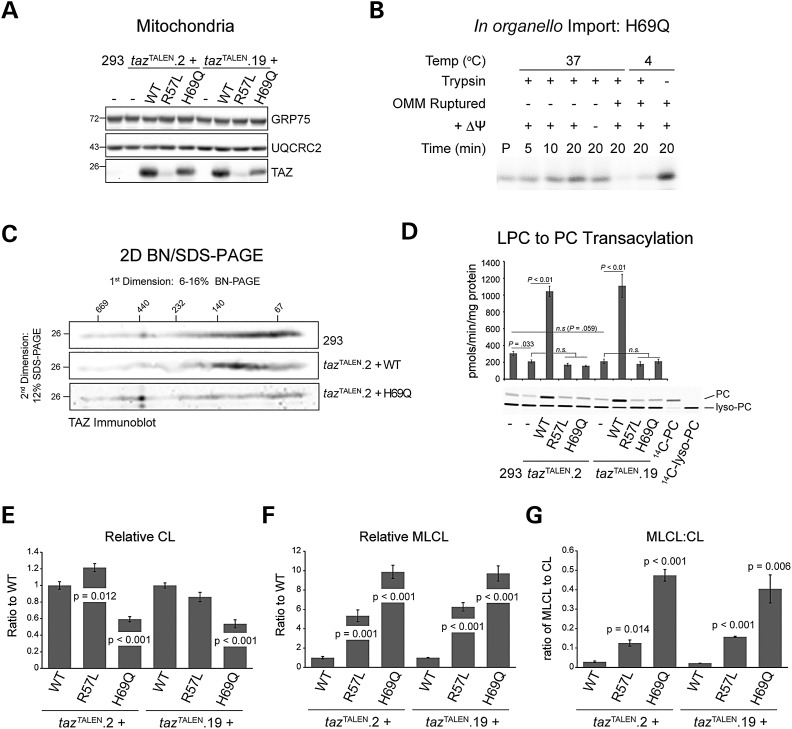
The X-linked disease Barth syndrome (BTHS) is caused by mutations in TAZ; TAZ is the main determinant of the final acyl chain composition of the mitochondrial-specific phospholipid, cardiolipin. To date, a detailed characterization of endogenous TAZ has only been performed in yeast. Further, why a given BTHS-associated missense mutation impairs TAZ function has only been determined in a yeast model of this human disease. Presently, the detailed characterization of yeast tafazzin harboring individual BTHS mutations at evolutionarily conserved residues has identified seven distinct loss-of-function mechanisms caused by patient-associated missense alleles. However, whether the biochemical consequences associated with individual mutations also occur in the context of human TAZ in a validated mammalian model has not been demonstrated. Here, utilizing newly established monoclonal antibodies capable of detecting endogenous TAZ, we demonstrate that mammalian TAZ, like its yeast counterpart, is localized to the mitochondrion where it adopts an extremely protease-resistant fold, associates non-integrally with intermembrane space-facing membranes and assembles in a range of complexes. Even though multiple isoforms are expressed at the mRNA level, only a single polypeptide that co-migrates with the human isoform lacking exon 5 is expressed in human skin fibroblasts, HEK293 cells, and murine heart and liver mitochondria. Finally, using a new genome-edited mammalian BTHS cell culture model, we demonstrate that the loss-of-function mechanisms for two BTHS alleles that represent two of the seven functional classes of BTHS mutation as originally defined in yeast, are the same when modeled in human TAZ.
© The Author 2016. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures

Similar articles
-
Tafazzin deficiency impairs CoA-dependent oxidative metabolism in cardiac mitochondria.J Biol Chem. 2020 Aug 28;295(35):12485-12497. doi: 10.1074/jbc.RA119.011229. Epub 2020 Jul 14. J Biol Chem. 2020. PMID: 32665401 Free PMC article.
-
Barth syndrome mutations that cause tafazzin complex lability.J Cell Biol. 2011 Feb 7;192(3):447-62. doi: 10.1083/jcb.201008177. J Cell Biol. 2011. PMID: 21300850 Free PMC article.
-
Increased mtDNA Abundance and Improved Function in Human Barth Syndrome Patient Fibroblasts Following AAV-TAZ Gene Delivery.Int J Mol Sci. 2019 Jul 11;20(14):3416. doi: 10.3390/ijms20143416. Int J Mol Sci. 2019. PMID: 31336787 Free PMC article.
-
Cardiolipin function in the yeast S. cerevisiae and the lessons learned for Barth syndrome.J Inherit Metab Dis. 2022 Jan;45(1):60-71. doi: 10.1002/jimd.12447. Epub 2021 Oct 19. J Inherit Metab Dis. 2022. PMID: 34626131 Free PMC article. Review.
-
Mitochondrial dysfunctions in barth syndrome.IUBMB Life. 2019 Jul;71(7):791-801. doi: 10.1002/iub.2018. Epub 2019 Feb 11. IUBMB Life. 2019. PMID: 30746873 Free PMC article. Review.
Cited by
-
Functional diversity among cardiolipin binding sites on the mitochondrial ADP/ATP carrier.EMBO J. 2024 Jul;43(14):2979-3008. doi: 10.1038/s44318-024-00132-2. Epub 2024 Jun 5. EMBO J. 2024. PMID: 38839991 Free PMC article.
-
Mechanism for Remodeling of the Acyl Chain Composition of Cardiolipin Catalyzed by Saccharomyces cerevisiae Tafazzin.J Biol Chem. 2016 Jul 22;291(30):15491-502. doi: 10.1074/jbc.M116.718510. Epub 2016 Jun 6. J Biol Chem. 2016. PMID: 27268057 Free PMC article.
-
Small heat shock proteins operate as molecular chaperones in the mitochondrial intermembrane space.Nat Cell Biol. 2023 Mar;25(3):467-480. doi: 10.1038/s41556-022-01074-9. Epub 2023 Jan 23. Nat Cell Biol. 2023. PMID: 36690850 Free PMC article.
-
Proteolytic Control of Lipid Metabolism.ACS Chem Biol. 2019 Nov 15;14(11):2406-2423. doi: 10.1021/acschembio.9b00695. Epub 2019 Sep 30. ACS Chem Biol. 2019. PMID: 31503446 Free PMC article. Review.
-
Cardiolipin coordinates inflammatory metabolic reprogramming through regulation of Complex II disassembly and degradation.Sci Adv. 2023 Feb 3;9(5):eade8701. doi: 10.1126/sciadv.ade8701. Epub 2023 Feb 3. Sci Adv. 2023. PMID: 36735777 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
