

Early-Life Social Isolation Stress Increases Kappa Opioid Receptor Responsiveness and Downregulates the Dopamine System
- PMID: 26860203
- PMCID: PMC4946054
- DOI: 10.1038/npp.2016.21
Early-Life Social Isolation Stress Increases Kappa Opioid Receptor Responsiveness and Downregulates the Dopamine System
Abstract
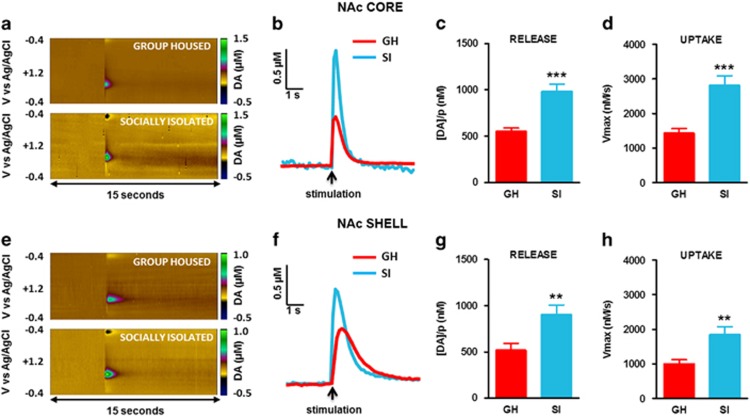
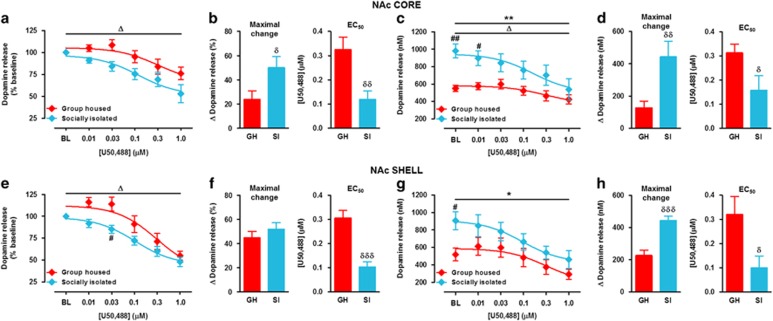
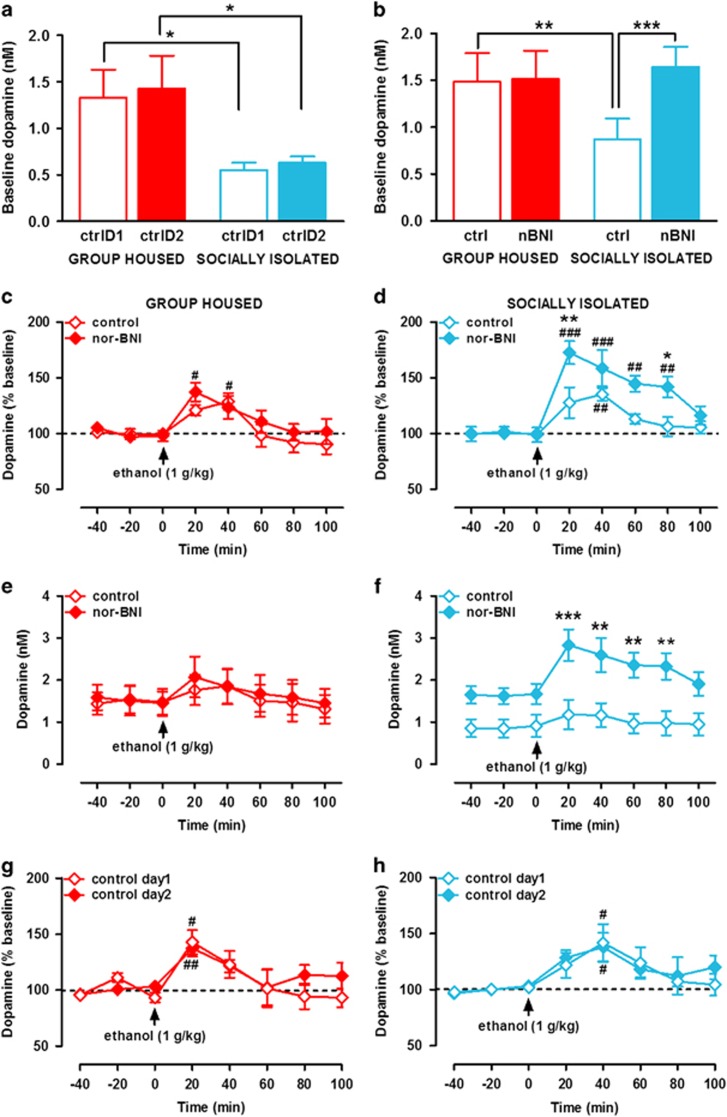
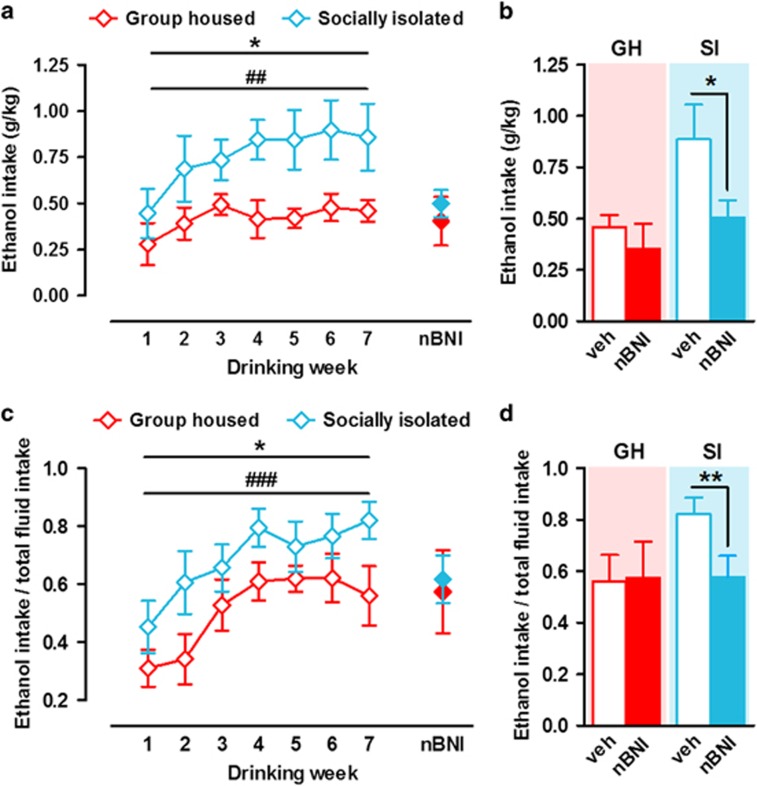
Chronic early-life stress increases vulnerability to alcoholism and anxiety disorders during adulthood. Similarly, rats reared in social isolation (SI) during adolescence exhibit augmented ethanol intake and anxiety-like behaviors compared with group housed (GH) rats. Prior studies suggest that disruption of dopamine (DA) signaling contributes to SI-associated behaviors, although the mechanisms underlying these alterations are not fully understood. Kappa opioid receptors (KORs) have an important role in regulating mesolimbic DA signaling, and other kinds of stressors have been shown to augment KOR function. Therefore, we tested the hypothesis that SI-induced increases in KOR function contribute to the dysregulation of NAc DA and the escalation in ethanol intake associated with SI. Our ex vivo voltammetry experiments showed that the inhibitory effects of the kappa agonist U50,488 on DA release were significantly enhanced in the NAc core and shell of SI rats. Dynorphin levels in NAc tissue were observed to be lower in SI rats. Microdialysis in freely moving rats revealed that SI was also associated with reduced baseline DA levels, and pretreatment with the KOR antagonist nor-binaltorphimine (nor-BNI) increased DA levels selectively in SI subjects. Acute ethanol elevated DA in SI and GH rats and nor-BNI pretreatment augmented this effect in SI subjects, while having no effect on ethanol-stimulated DA release in GH rats. Together, these data suggest that KORs may have increased responsiveness following SI, which could lead to hypodopaminergia and contribute to an increased drive to consume ethanol. Indeed, SI rats exhibited greater ethanol intake and preference and KOR blockade selectively attenuated ethanol intake in SI rats. Collectively, the findings that nor-BNI reversed SI-mediated hypodopaminergic state and escalated ethanol intake suggest that KOR antagonists may represent a promising therapeutic strategy for the treatment of alcohol use disorders, particularly in cases linked to chronic early-life stress.
Figures
Similar articles
-
Social isolation rearing increases nucleus accumbens dopamine and norepinephrine responses to acute ethanol in adulthood.Alcohol Clin Exp Res. 2014 Nov;38(11):2770-9. doi: 10.1111/acer.12555. Alcohol Clin Exp Res. 2014. PMID: 25421514 Free PMC article.
-
Distinct Effects of Nalmefene on Dopamine Uptake Rates and Kappa Opioid Receptor Activity in the Nucleus Accumbens Following Chronic Intermittent Ethanol Exposure.Int J Mol Sci. 2016 Jul 27;17(8):1216. doi: 10.3390/ijms17081216. Int J Mol Sci. 2016. PMID: 27472317 Free PMC article.
-
Switch from excitatory to inhibitory actions of ethanol on dopamine levels after chronic exposure: Role of kappa opioid receptors.Neuropharmacology. 2016 Nov;110(Pt A):190-197. doi: 10.1016/j.neuropharm.2016.07.022. Epub 2016 Jul 20. Neuropharmacology. 2016. PMID: 27450094 Free PMC article.
-
Dynorphin/Kappa Opioid Receptor Signaling in Preclinical Models of Alcohol, Drug, and Food Addiction.Int Rev Neurobiol. 2017;136:53-88. doi: 10.1016/bs.irn.2017.08.001. Epub 2017 Sep 12. Int Rev Neurobiol. 2017. PMID: 29056156 Review.
-
Role of the Dynorphin/Kappa Opioid Receptor System in the Motivational Effects of Ethanol.Alcohol Clin Exp Res. 2017 Aug;41(8):1402-1418. doi: 10.1111/acer.13406. Epub 2017 Jun 5. Alcohol Clin Exp Res. 2017. PMID: 28425121 Free PMC article. Review.
Cited by
-
Early Life Stress and Risks for Opioid Misuse: Review of Data Supporting Neurobiological Underpinnings.J Pers Med. 2021 Apr 19;11(4):315. doi: 10.3390/jpm11040315. J Pers Med. 2021. PMID: 33921642 Free PMC article. Review.
-
Adolescent Social Isolation as a Model of Heightened Vulnerability to Comorbid Alcoholism and Anxiety Disorders.Alcohol Clin Exp Res. 2016 Jun;40(6):1202-14. doi: 10.1111/acer.13075. Epub 2016 May 7. Alcohol Clin Exp Res. 2016. PMID: 27154240 Free PMC article. Review.
-
Stress-Induced Changes in the Endogenous Opioid System Cause Dysfunction of Pain and Emotion Regulation.Int J Mol Sci. 2023 Jul 20;24(14):11713. doi: 10.3390/ijms241411713. Int J Mol Sci. 2023. PMID: 37511469 Free PMC article. Review.
-
Sex differences in ethanol consumption and drinking despite negative consequences following adolescent social isolation stress in male and female rats.Physiol Behav. 2023 Nov 1;271:114322. doi: 10.1016/j.physbeh.2023.114322. Epub 2023 Aug 12. Physiol Behav. 2023. PMID: 37573960 Free PMC article.
-
Stress-Induced Enhancement of Ethanol Intake in C57BL/6J Mice with a History of Chronic Ethanol Exposure: Involvement of Kappa Opioid Receptors.Front Cell Neurosci. 2016 Feb 23;10:45. doi: 10.3389/fncel.2016.00045. eCollection 2016. Front Cell Neurosci. 2016. PMID: 26941607 Free PMC article.
References
-
- Anda RF, Whitfield CL, Felitti VJ, Chapman D, Edwards VJ, Dube SR et al (2002). Adverse childhood experiences, alcoholic parents, and later risk of alcoholism and depression. Psychiatr Serv 53: 1001–1009. - PubMed
-
- Beardsley PM, Howard JL, Shelton KL, Carroll FI (2005). Differential effects of the novel kappa opioid receptor antagonist, JDTic, on reinstatement of cocaine-seeking induced by footshock stressors vs cocaine primes and its antidepressant-like effects in rats. Psychopharmacology 183: 118–126. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
