

Central Gain Restores Auditory Processing following Near-Complete Cochlear Denervation
- PMID: 26833137
- PMCID: PMC4760846
- DOI: 10.1016/j.neuron.2015.12.041
Central Gain Restores Auditory Processing following Near-Complete Cochlear Denervation
Abstract
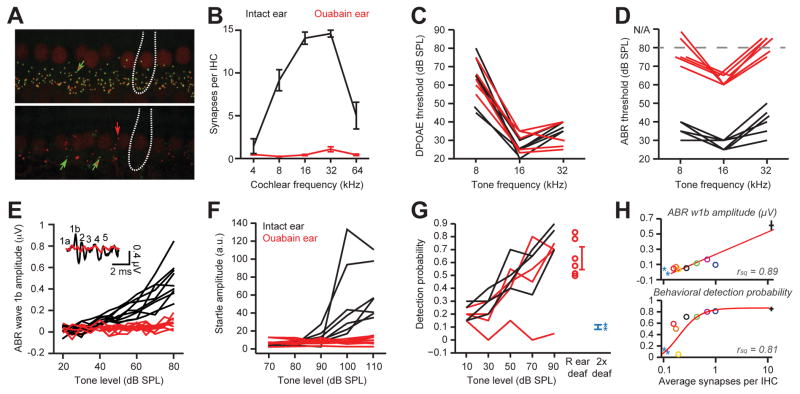
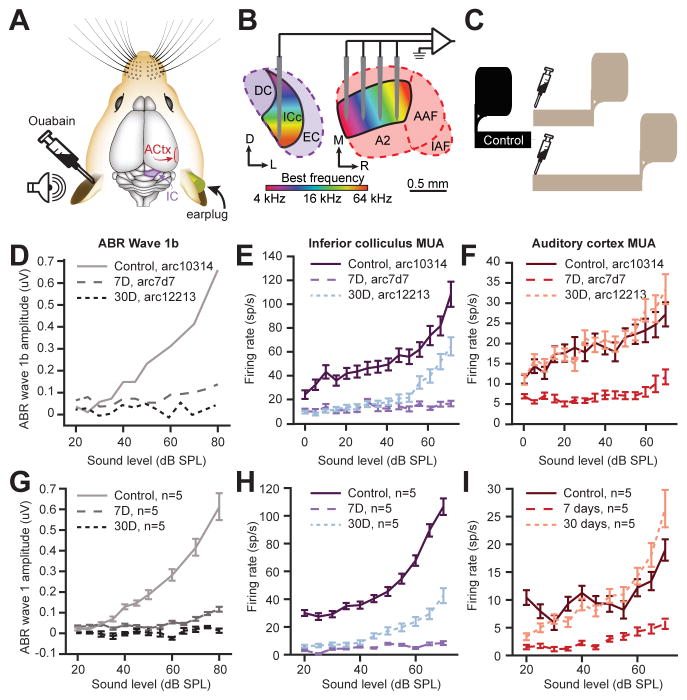
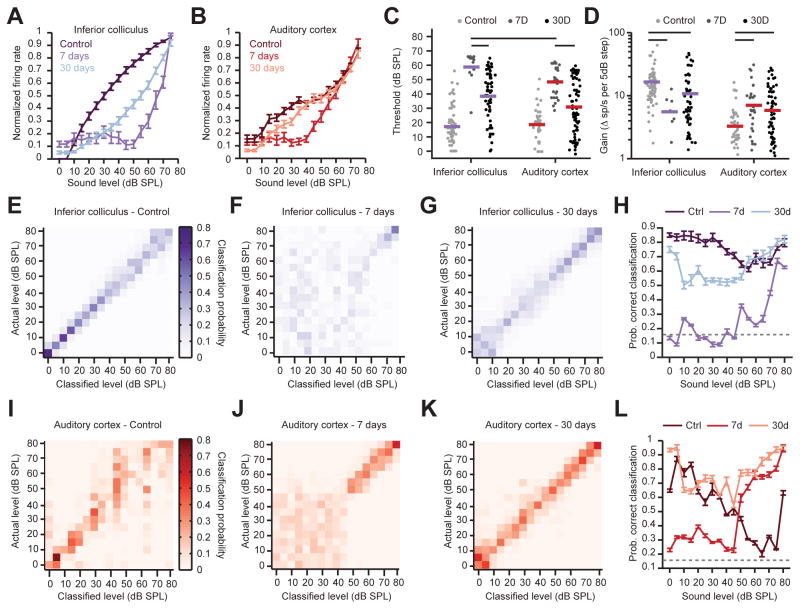
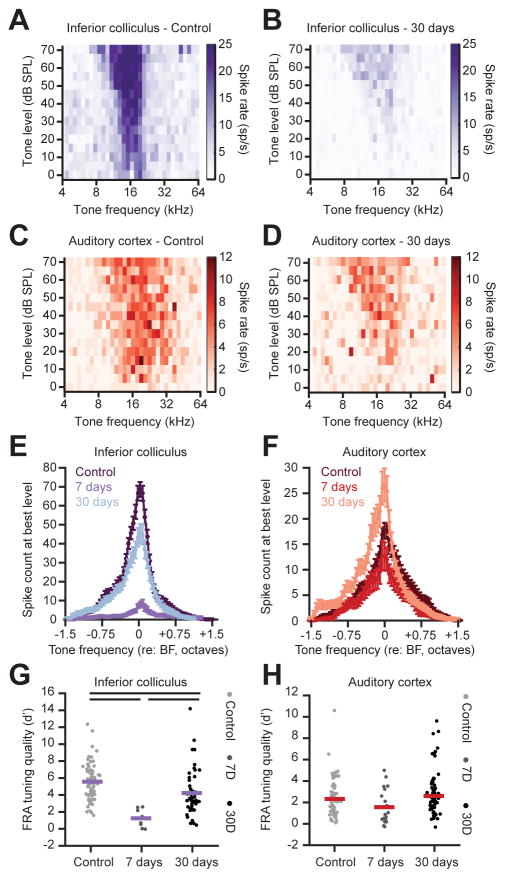
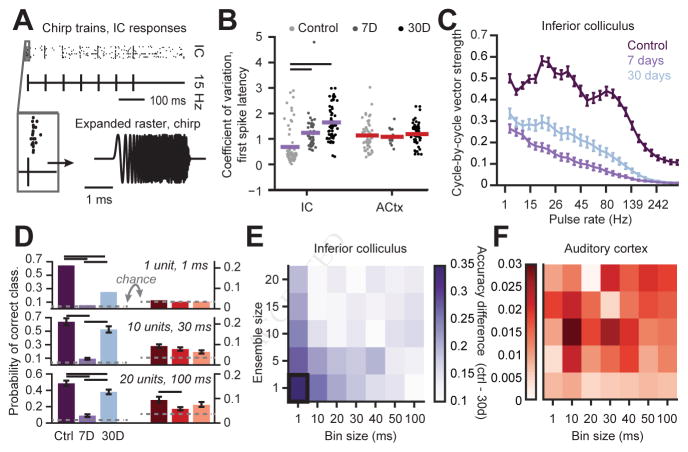
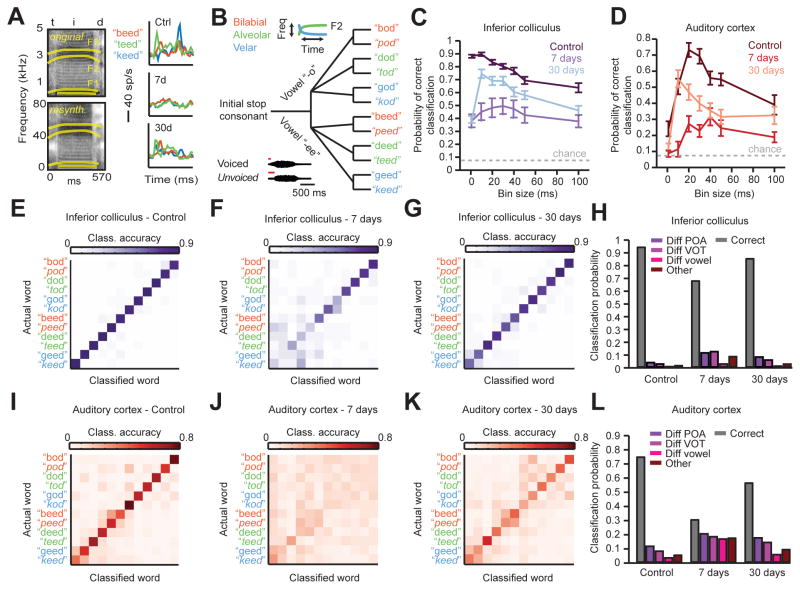
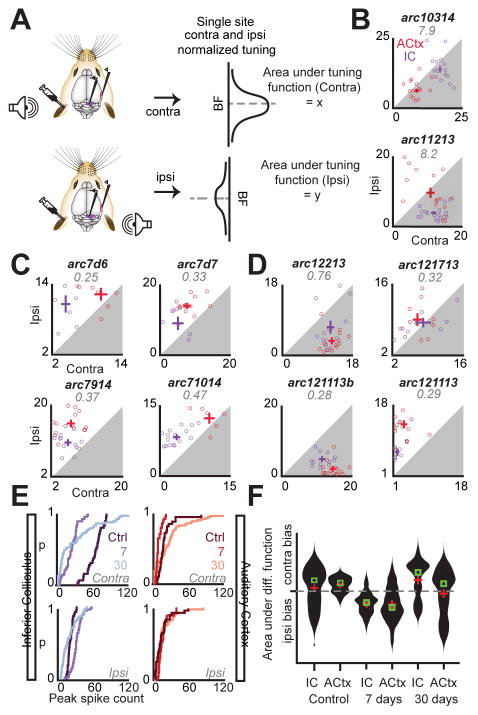
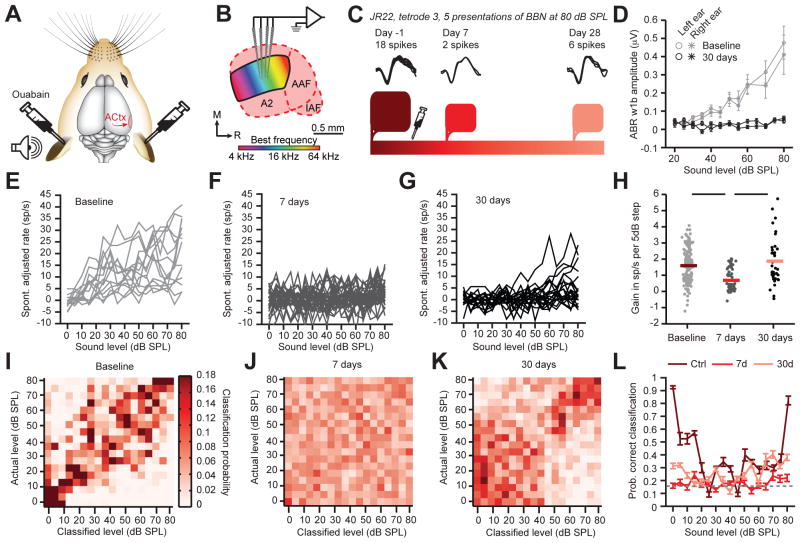
Sensory organ damage induces a host of cellular and physiological changes in the periphery and the brain. Here, we show that some aspects of auditory processing recover after profound cochlear denervation due to a progressive, compensatory plasticity at higher stages of the central auditory pathway. Lesioning >95% of cochlear nerve afferent synapses, while sparing hair cells, in adult mice virtually eliminated the auditory brainstem response and acoustic startle reflex, yet tone detection behavior was nearly normal. As sound-evoked responses from the auditory nerve grew progressively weaker following denervation, sound-evoked activity in the cortex-and, to a lesser extent, the midbrain-rebounded or surpassed control levels. Increased central gain supported the recovery of rudimentary sound features encoded by firing rate, but not features encoded by precise spike timing such as modulated noise or speech. These findings underscore the importance of central plasticity in the perceptual sequelae of cochlear hearing impairment.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
Comment in
-
The Ying and Yang of Auditory Nerve Damage.Neuron. 2016 Feb 17;89(4):680-2. doi: 10.1016/j.neuron.2016.02.007. Neuron. 2016. PMID: 26889808
Similar articles
-
The Ying and Yang of Auditory Nerve Damage.Neuron. 2016 Feb 17;89(4):680-2. doi: 10.1016/j.neuron.2016.02.007. Neuron. 2016. PMID: 26889808
-
Synaptopathy in the Aging Cochlea: Characterizing Early-Neural Deficits in Auditory Temporal Envelope Processing.J Neurosci. 2018 Aug 8;38(32):7108-7119. doi: 10.1523/JNEUROSCI.3240-17.2018. Epub 2018 Jul 5. J Neurosci. 2018. PMID: 29976623 Free PMC article.
-
Persistent Thalamic Sound Processing Despite Profound Cochlear Denervation.Front Neural Circuits. 2016 Aug 31;10:72. doi: 10.3389/fncir.2016.00072. eCollection 2016. Front Neural Circuits. 2016. PMID: 27630546 Free PMC article.
-
Audibility, speech perception and processing of temporal cues in ribbon synaptic disorders due to OTOF mutations.Hear Res. 2015 Dec;330(Pt B):200-12. doi: 10.1016/j.heares.2015.07.007. Epub 2015 Jul 15. Hear Res. 2015. PMID: 26188103 Review.
-
Central nervous system plasticity during hair cell loss and regeneration.Hear Res. 2000 Sep;147(1-2):145-59. doi: 10.1016/s0378-5955(00)00128-3. Hear Res. 2000. PMID: 10962181 Review.
Cited by
-
Objective evidence of temporal processing deficits in older adults.Hear Res. 2020 Nov;397:108053. doi: 10.1016/j.heares.2020.108053. Epub 2020 Aug 16. Hear Res. 2020. PMID: 32863099 Free PMC article. Review.
-
Primary Neural Degeneration in Noise-Exposed Human Cochleas: Correlations with Outer Hair Cell Loss and Word-Discrimination Scores.J Neurosci. 2021 May 19;41(20):4439-4447. doi: 10.1523/JNEUROSCI.3238-20.2021. Epub 2021 Apr 21. J Neurosci. 2021. PMID: 33883202 Free PMC article.
-
Auditory-frontal Channeling in α and β Bands is Altered by Age-related Hearing Loss and Relates to Speech Perception in Noise.Neuroscience. 2019 Dec 15;423:18-28. doi: 10.1016/j.neuroscience.2019.10.044. Epub 2019 Nov 6. Neuroscience. 2019. PMID: 31705894 Free PMC article.
-
Hidden hearing loss selectively impairs neural adaptation to loud sound environments.Nat Commun. 2018 Oct 16;9(1):4298. doi: 10.1038/s41467-018-06777-y. Nat Commun. 2018. PMID: 30327471 Free PMC article.
-
Age-related hearing loss is associated with alterations in temporal envelope processing in different neural generators along the auditory pathway.Front Neurol. 2022 Aug 5;13:905017. doi: 10.3389/fneur.2022.905017. eCollection 2022. Front Neurol. 2022. PMID: 35989932 Free PMC article.
References
-
- Berlin CI, Hood LJ, Morlet T, Wilensky D, Li L, Mattingly KR, Taylor-Jeanfreau J, Keats BJB, John PS, Montgomery E, et al. Multi-site diagnosis and management of 260 patients with Auditory Neuropathy/Dys-synchrony (Auditory Neuropathy Spectrum Disorder) Int J Audiol. 2010;49:30–43. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
