

Affinity and competition for TBP are molecular determinants of gene expression noise
- PMID: 26832815
- PMCID: PMC4740812
- DOI: 10.1038/ncomms10417
Affinity and competition for TBP are molecular determinants of gene expression noise
Abstract
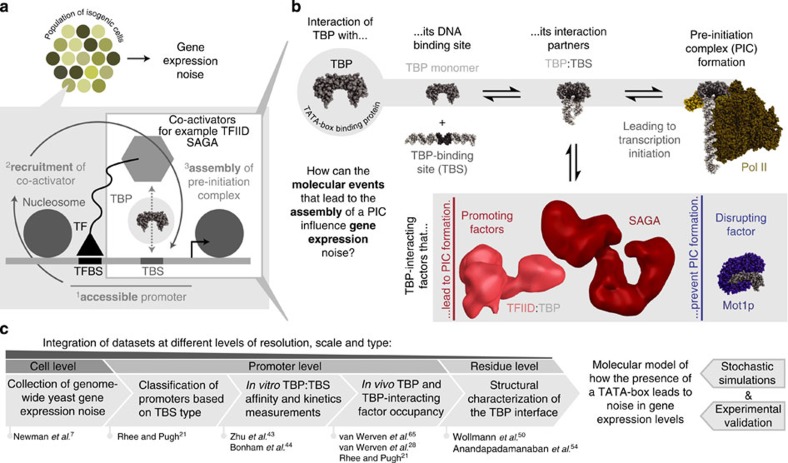
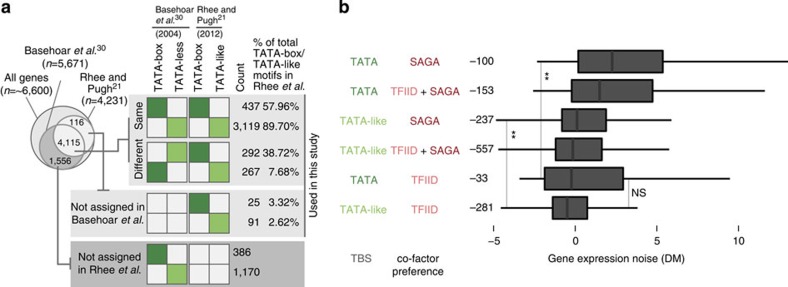
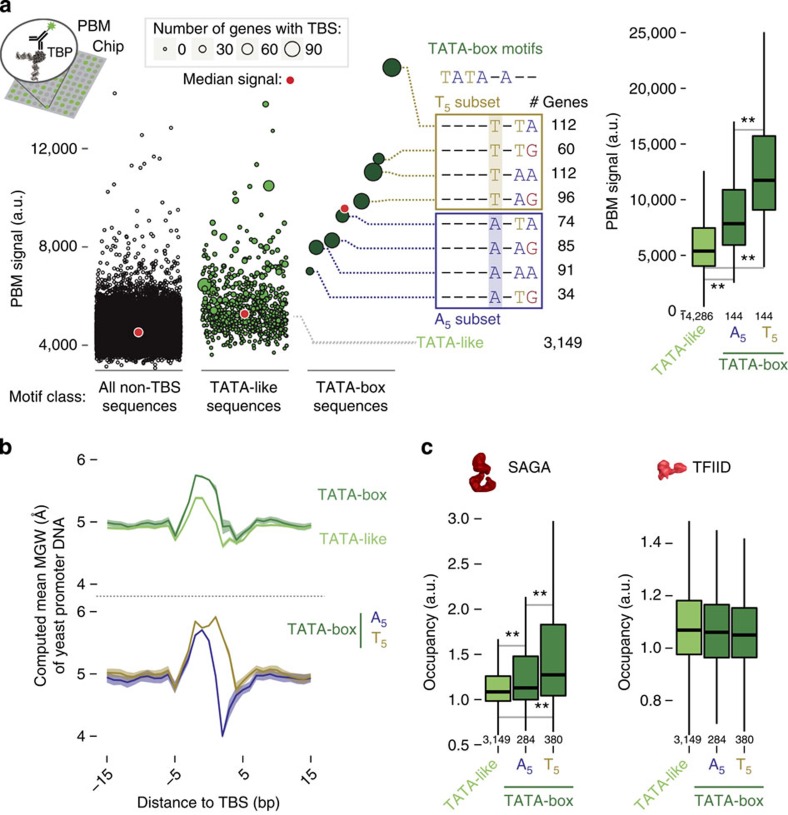
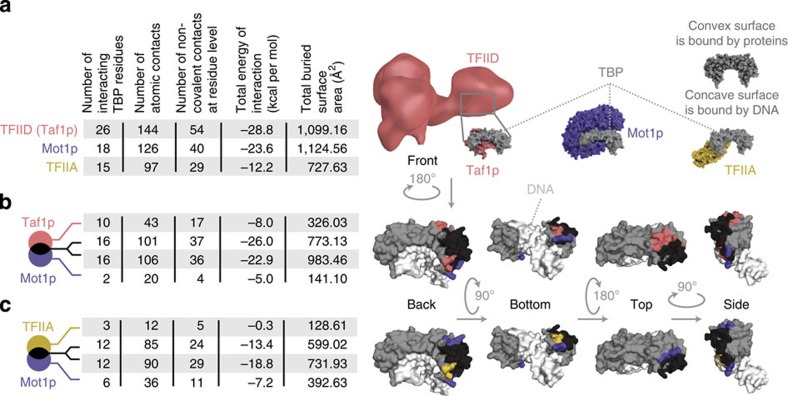
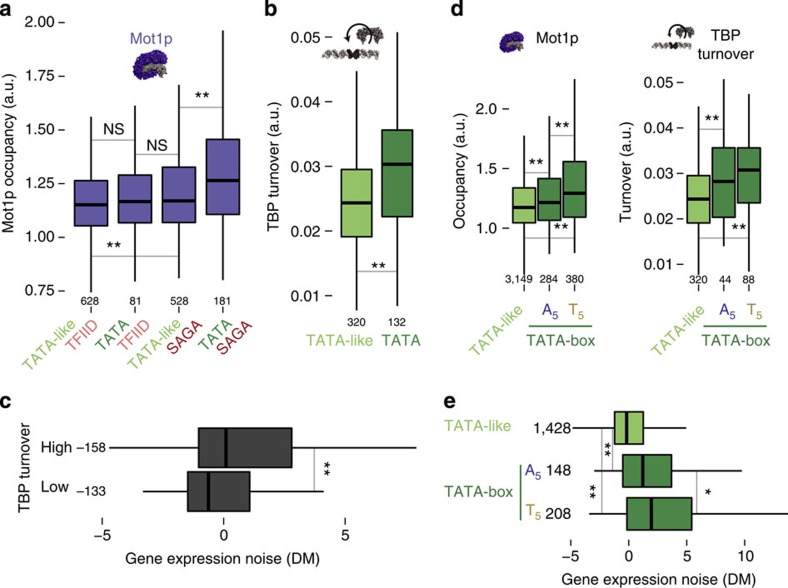
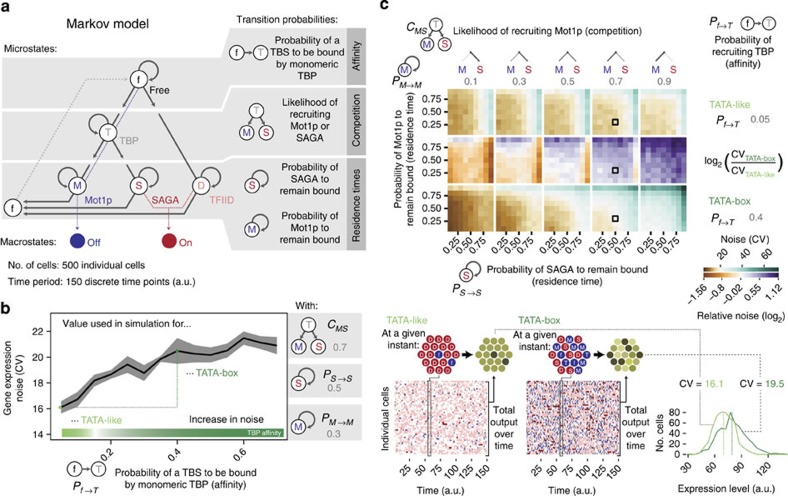
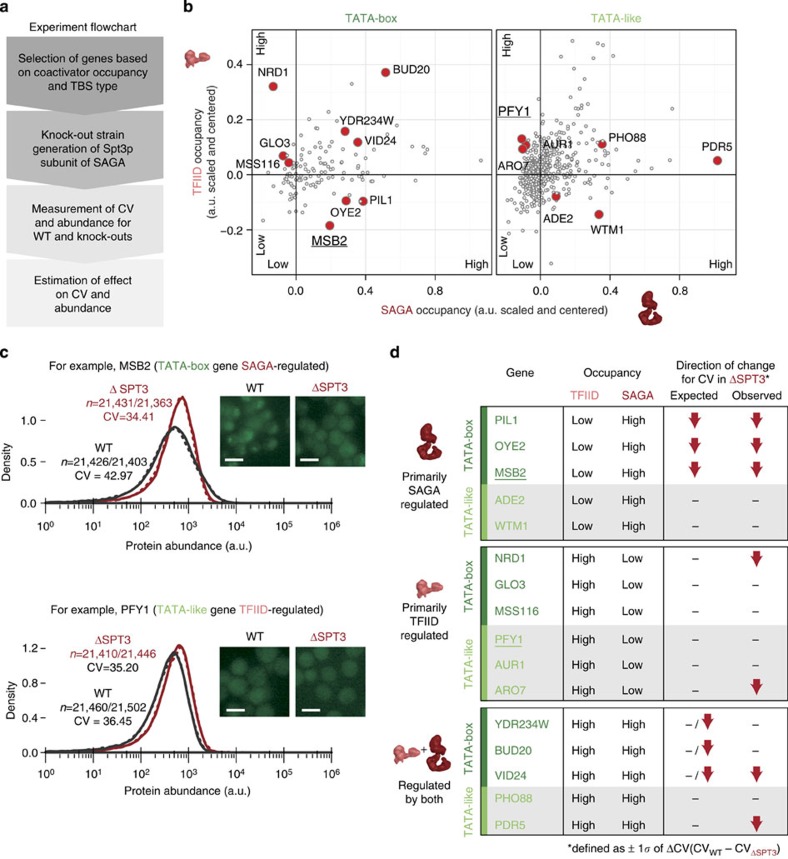
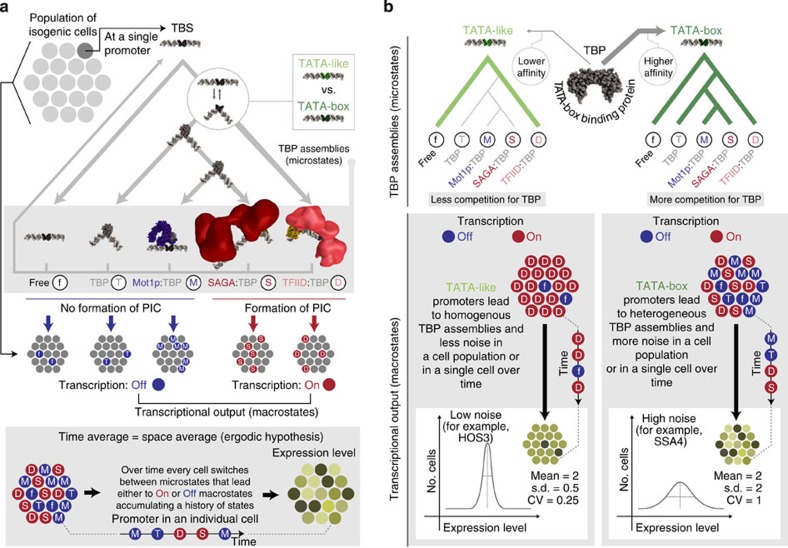
Cell-to-cell variation in gene expression levels (noise) generates phenotypic diversity and is an important phenomenon in evolution, development and disease. TATA-box binding protein (TBP) is an essential factor that is required at virtually every eukaryotic promoter to initiate transcription. While the presence of a TATA-box motif in the promoter has been strongly linked with noise, the molecular mechanism driving this relationship is less well understood. Through an integrated analysis of multiple large-scale data sets, computer simulation and experimental validation in yeast, we provide molecular insights into how noise arises as an emergent property of variable binding affinity of TBP for different promoter sequences, competition between interaction partners to bind the same surface on TBP (to either promote or disrupt transcription initiation) and variable residence times of TBP complexes at a promoter. These determinants may be fine-tuned under different conditions and during evolution to modulate eukaryotic gene expression noise.
Figures
Similar articles
-
Global role of TATA box-binding protein recruitment to promoters in mediating gene expression profiles.Mol Cell Biol. 2004 Sep;24(18):8104-12. doi: 10.1128/MCB.24.18.8104-8112.2004. Mol Cell Biol. 2004. PMID: 15340072 Free PMC article.
-
Sequence-dependent solution structure and motions of 13 TATA/TBP (TATA-box binding protein) complexes.Biopolymers. 2003 Jun;69(2):216-43. doi: 10.1002/bip.10409. Biopolymers. 2003. PMID: 12767124
-
Structural and functional characterization on the interaction of yeast TFIID subunit TAF1 with TATA-binding protein.J Mol Biol. 2004 Jun 11;339(4):681-93. doi: 10.1016/j.jmb.2004.04.020. J Mol Biol. 2004. PMID: 15165843
-
X-ray crystallographic studies of eukaryotic transcription initiation factors.Philos Trans R Soc Lond B Biol Sci. 1996 Apr 29;351(1339):483-9. doi: 10.1098/rstb.1996.0046. Philos Trans R Soc Lond B Biol Sci. 1996. PMID: 8735270 Review.
-
TBP homologues in embryo transcription: who does what?EMBO Rep. 2007 Nov;8(11):1016-8. doi: 10.1038/sj.embor.7401093. EMBO Rep. 2007. PMID: 17972900 Free PMC article. Review. No abstract available.
Cited by
-
Structure of SAGA and mechanism of TBP deposition on gene promoters.Nature. 2020 Jan;577(7792):711-716. doi: 10.1038/s41586-020-1944-2. Epub 2020 Jan 22. Nature. 2020. PMID: 31969704
-
The switch of DNA states filtering the extrinsic noise in the system of frequency modulation.Sci Rep. 2021 Aug 11;11(1):16309. doi: 10.1038/s41598-021-95365-0. Sci Rep. 2021. PMID: 34381062 Free PMC article.
-
Two promoters integrate multiple enhancer inputs to drive wild-type knirps expression in the Drosophila melanogaster embryo.Genetics. 2021 Dec 10;219(4):iyab154. doi: 10.1093/genetics/iyab154. Genetics. 2021. PMID: 34849867 Free PMC article.
-
DNA-induced spatial entrapment of general transcription machinery can stabilize gene expression in a nondividing cell.Proc Natl Acad Sci U S A. 2022 Jan 25;119(4):e2116091119. doi: 10.1073/pnas.2116091119. Proc Natl Acad Sci U S A. 2022. PMID: 35074915 Free PMC article.
-
Comparative studies of genomic and epigenetic factors influencing transcriptional variation in two insect species.G3 (Bethesda). 2022 Nov 4;12(11):jkac230. doi: 10.1093/g3journal/jkac230. G3 (Bethesda). 2022. PMID: 36137211 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
