

N(6)-methyladenosine in mRNA disrupts tRNA selection and translation-elongation dynamics
- PMID: 26751643
- PMCID: PMC4826618
- DOI: 10.1038/nsmb.3148
N(6)-methyladenosine in mRNA disrupts tRNA selection and translation-elongation dynamics
Abstract
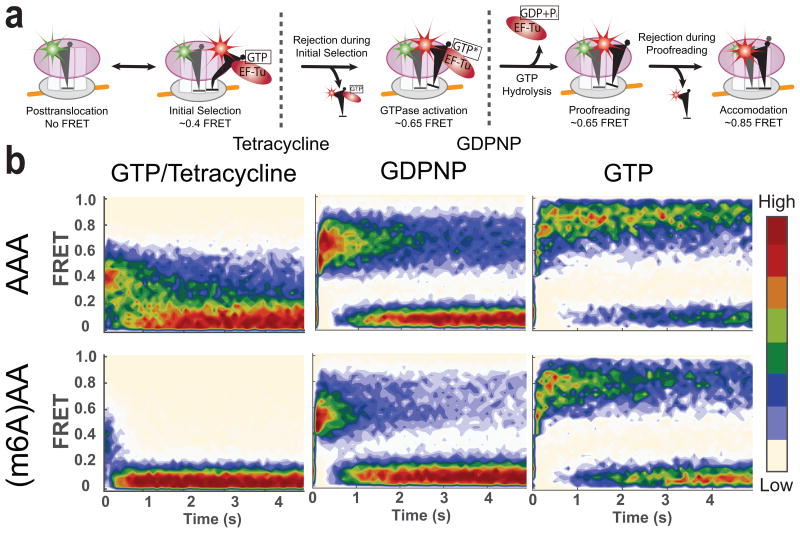
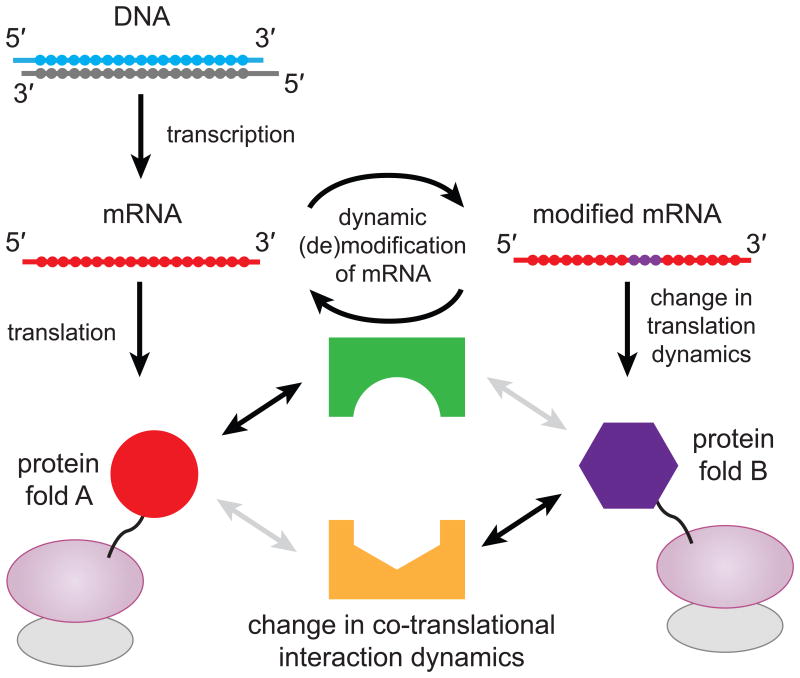
N(6)-methylation of adenosine (forming m(6)A) is the most abundant post-transcriptional modification within the coding region of mRNA, but its role during translation remains unknown. Here, we used bulk kinetic and single-molecule methods to probe the effect of m(6)A in mRNA decoding. Although m(6)A base-pairs with uridine during decoding, as shown by X-ray crystallographic analyses of Thermus thermophilus ribosomal complexes, our measurements in an Escherichia coli translation system revealed that m(6)A modification of mRNA acts as a barrier to tRNA accommodation and translation elongation. The interaction between an m(6)A-modified codon and cognate tRNA echoes the interaction between a near-cognate codon and tRNA, because delay in tRNA accommodation depends on the position and context of m(6)A within codons and on the accuracy level of translation. Overall, our results demonstrate that chemical modification of mRNA can change translational dynamics.
Figures
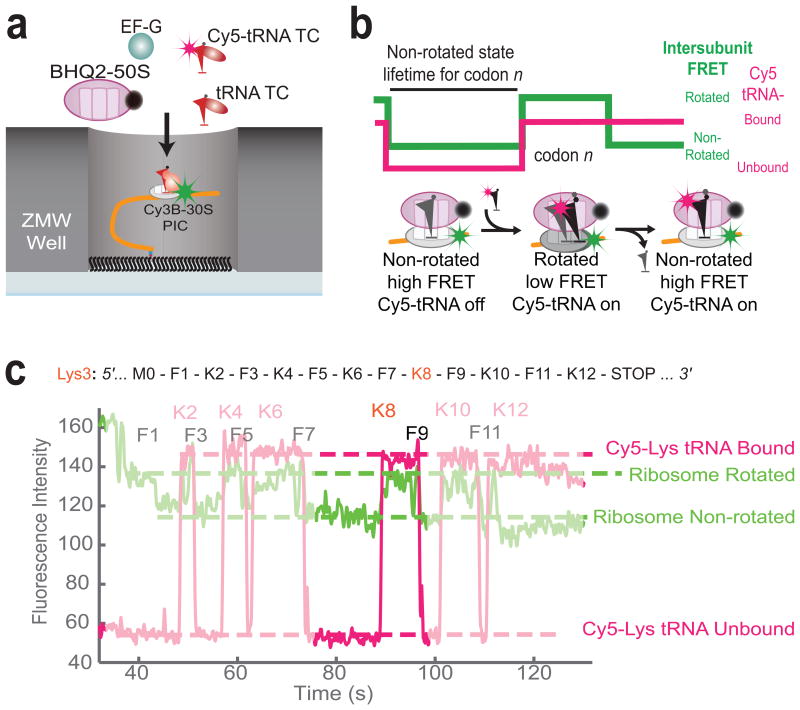
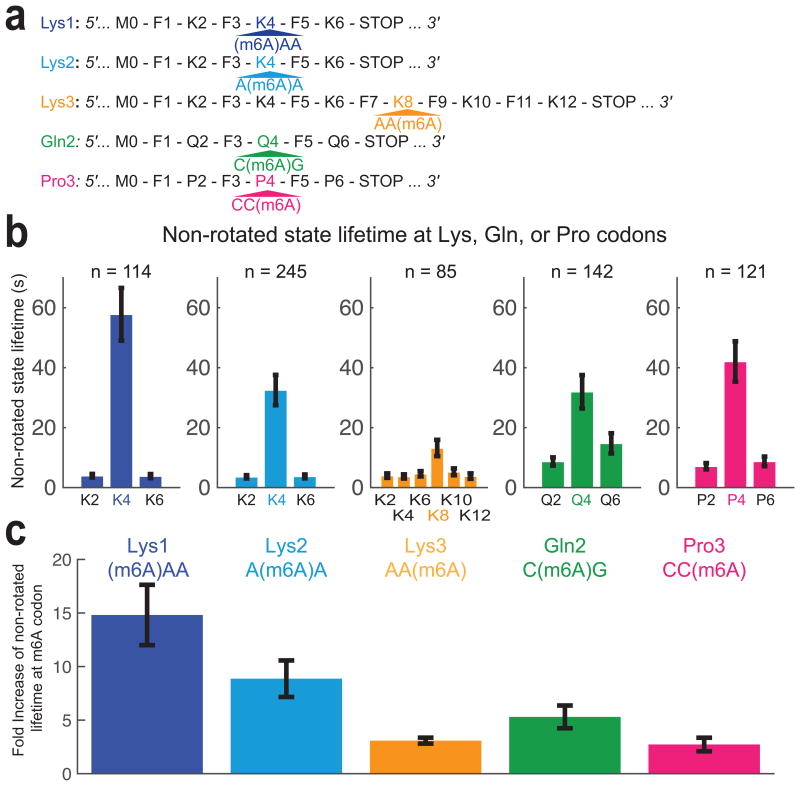
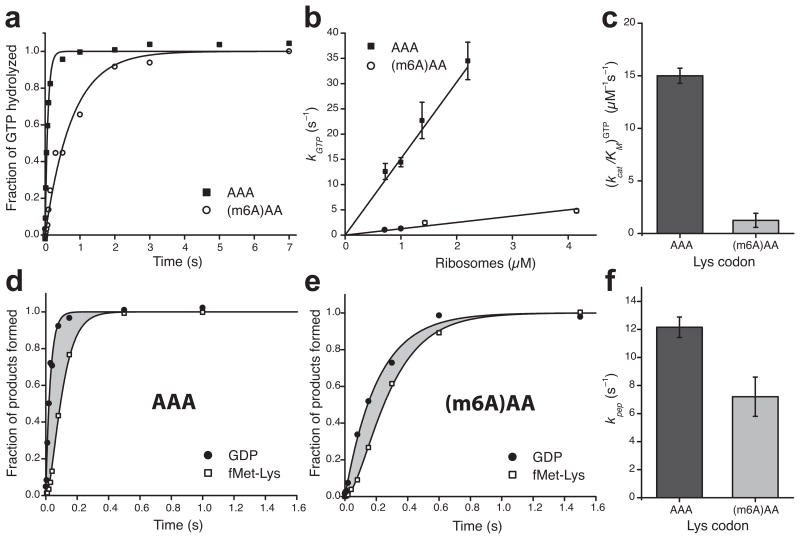
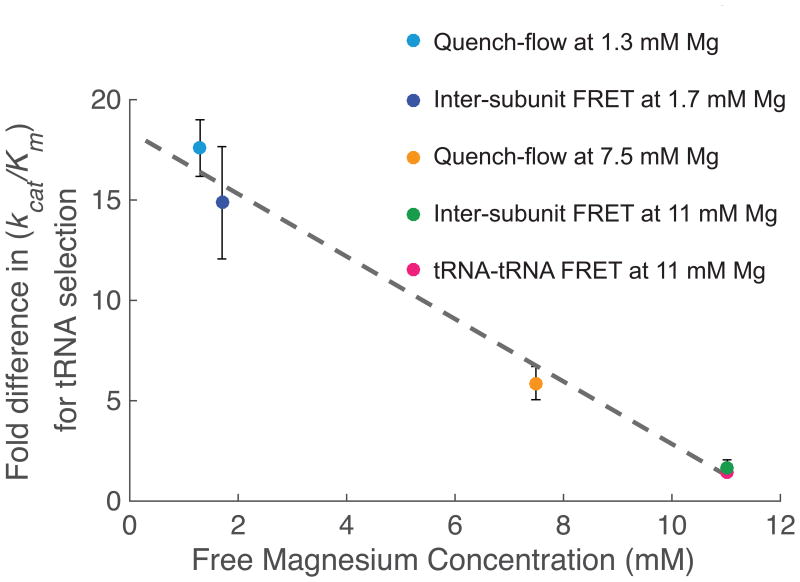
Similar articles
-
Tautomeric G•U pairs within the molecular ribosomal grip and fidelity of decoding in bacteria.Nucleic Acids Res. 2018 Aug 21;46(14):7425-7435. doi: 10.1093/nar/gky547. Nucleic Acids Res. 2018. PMID: 29931292 Free PMC article.
-
Pseudouridinylation of mRNA coding sequences alters translation.Proc Natl Acad Sci U S A. 2019 Nov 12;116(46):23068-23074. doi: 10.1073/pnas.1821754116. Epub 2019 Oct 31. Proc Natl Acad Sci U S A. 2019. PMID: 31672910 Free PMC article.
-
Disruption of evolutionarily correlated tRNA elements impairs accurate decoding.Proc Natl Acad Sci U S A. 2020 Jul 14;117(28):16333-16338. doi: 10.1073/pnas.2004170117. Epub 2020 Jun 29. Proc Natl Acad Sci U S A. 2020. PMID: 32601241 Free PMC article.
-
Structural insights into translational fidelity.Annu Rev Biochem. 2005;74:129-77. doi: 10.1146/annurev.biochem.74.061903.155440. Annu Rev Biochem. 2005. PMID: 15952884 Review.
-
Reversible RNA Modification N1-methyladenosine (m1A) in mRNA and tRNA.Genomics Proteomics Bioinformatics. 2018 Jun;16(3):155-161. doi: 10.1016/j.gpb.2018.03.003. Epub 2018 Jun 14. Genomics Proteomics Bioinformatics. 2018. PMID: 29908293 Free PMC article. Review.
Cited by
-
The epitranscriptome of Vero cells infected with SARS-CoV-2 assessed by direct RNA sequencing reveals m6A pattern changes and DRACH motif biases in viral and cellular RNAs.Front Cell Infect Microbiol. 2022 Aug 16;12:906578. doi: 10.3389/fcimb.2022.906578. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 36051243 Free PMC article.
-
Post-transcriptional gene regulation by mRNA modifications.Nat Rev Mol Cell Biol. 2017 Jan;18(1):31-42. doi: 10.1038/nrm.2016.132. Epub 2016 Nov 3. Nat Rev Mol Cell Biol. 2017. PMID: 27808276 Free PMC article. Review.
-
Specific RNA m6A modification sites in bone marrow mesenchymal stem cells from the jawbone marrow of type 2 diabetes patients with dental implant failure.Int J Oral Sci. 2023 Jan 12;15(1):6. doi: 10.1038/s41368-022-00202-3. Int J Oral Sci. 2023. PMID: 36631441 Free PMC article.
-
Eukaryotic Translation Elongation is Modulated by Single Natural Nucleotide Derivatives in the Coding Sequences of mRNAs.Genes (Basel). 2019 Jan 25;10(2):84. doi: 10.3390/genes10020084. Genes (Basel). 2019. PMID: 30691071 Free PMC article.
-
Relating Structure and Dynamics in RNA Biology.Cold Spring Harb Perspect Biol. 2019 Jul 1;11(7):a032474. doi: 10.1101/cshperspect.a032474. Cold Spring Harb Perspect Biol. 2019. PMID: 31262948 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
