

Exon-centric regulation of ATM expression is population-dependent and amenable to antisense modification by pseudoexon targeting
- PMID: 26732650
- PMCID: PMC4702124
- DOI: 10.1038/srep18741
Exon-centric regulation of ATM expression is population-dependent and amenable to antisense modification by pseudoexon targeting
Erratum in
-
Corrigendum: Exon-centric regulation of ATM expression is population-dependent and amenable to antisense modification by pseudoexon targeting.Sci Rep. 2016 May 9;6:25256. doi: 10.1038/srep25256. Sci Rep. 2016. PMID: 27157627 Free PMC article. No abstract available.
Abstract
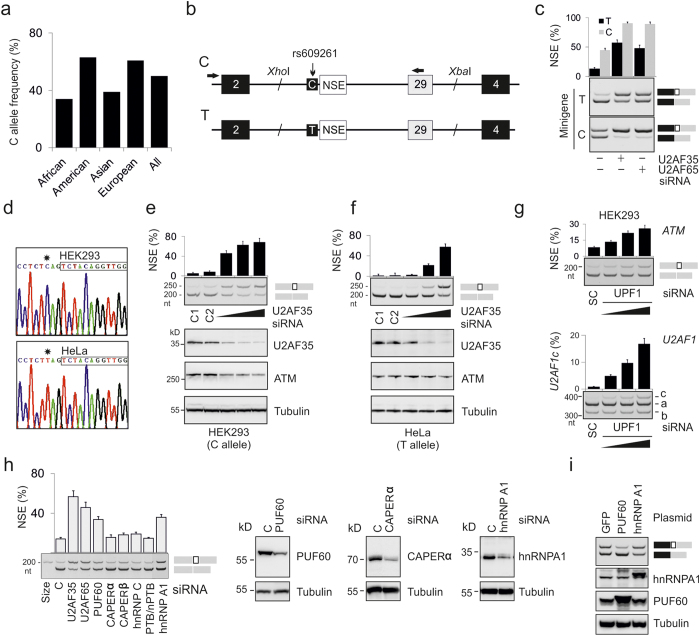
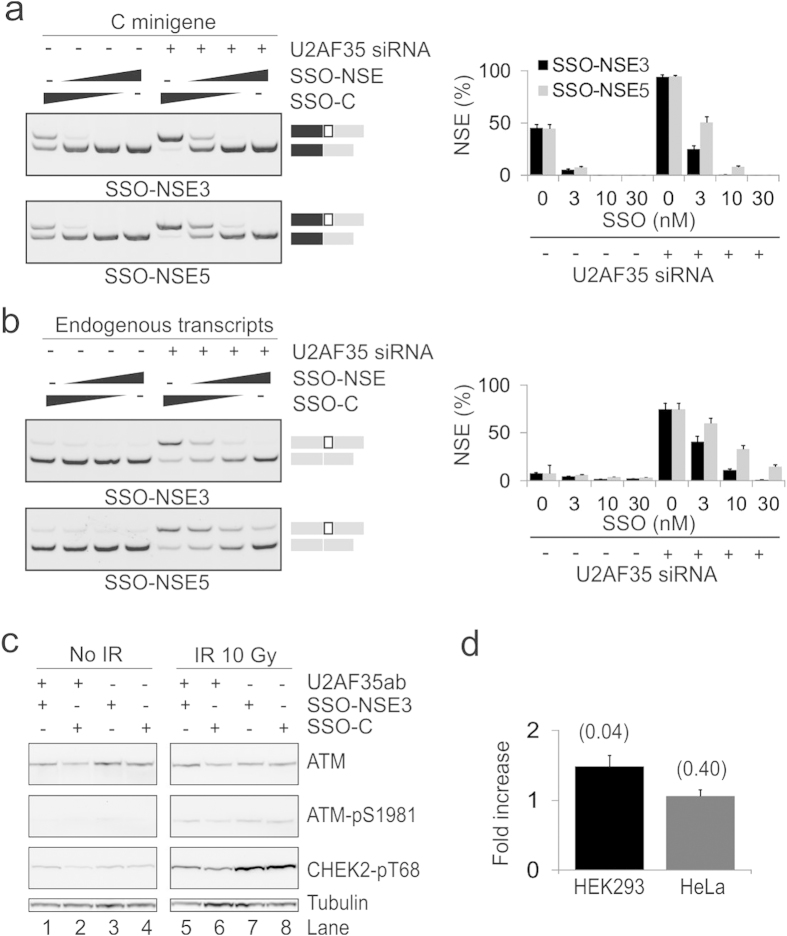
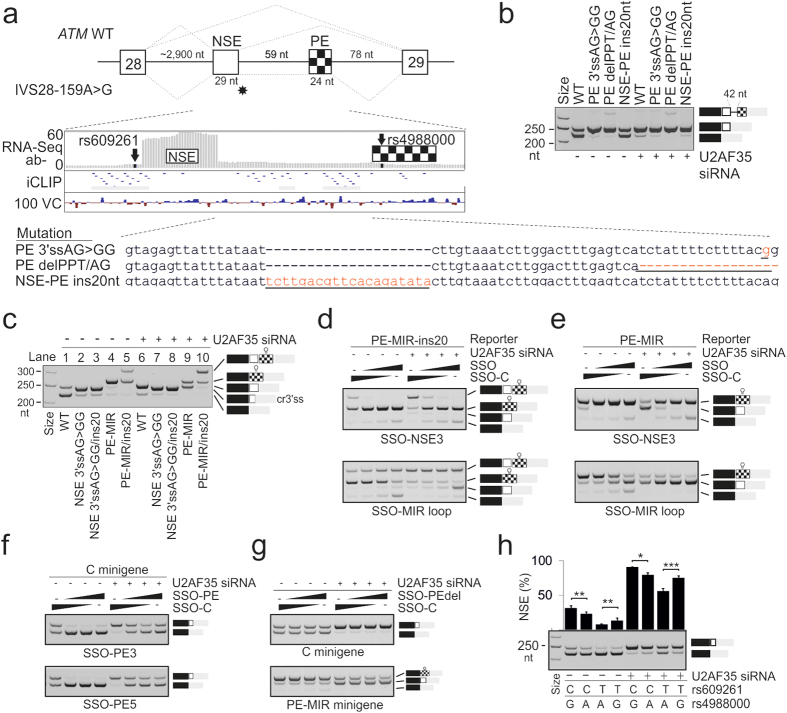
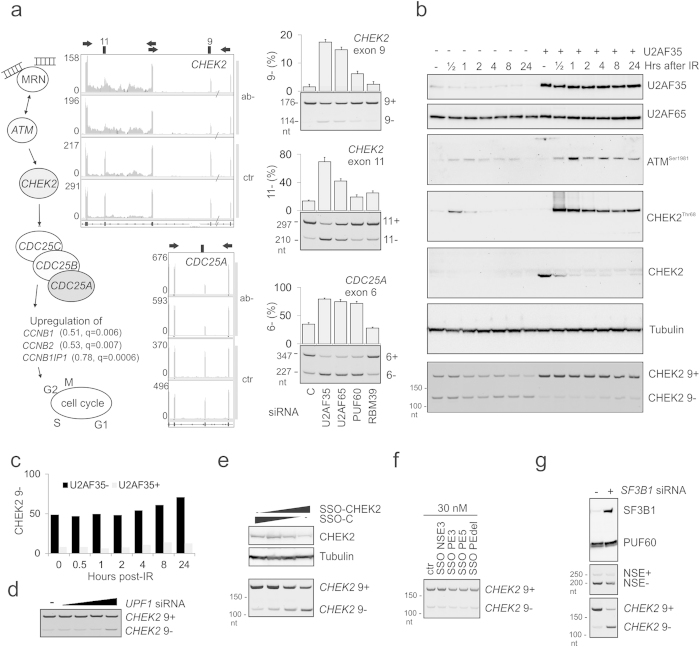
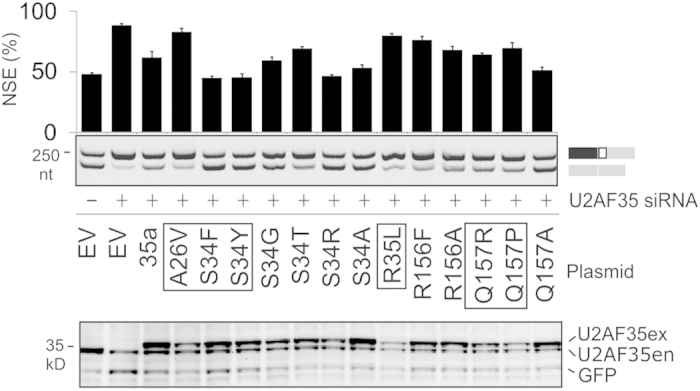
ATM is an important cancer susceptibility gene that encodes a critical apical kinase of the DNA damage response (DDR) pathway. We show that a key nonsense-mediated RNA decay switch exon (NSE) in ATM is repressed by U2AF, PUF60 and hnRNPA1. The NSE activation was haplotype-specific and was most promoted by cytosine at rs609621 in the NSE 3' splice-site (3'ss), which is predominant in high cancer risk populations. NSE levels were deregulated in leukemias and were influenced by the identity of U2AF35 residue 34. We also identify splice-switching oligonucleotides (SSOs) that exploit competition of adjacent pseudoexons to modulate NSE levels. The U2AF-regulated exon usage in the ATM signalling pathway was centred on the MRN/ATM-CHEK2-CDC25-cdc2/cyclin-B axis and preferentially involved transcripts implicated in cancer-associated gene fusions and chromosomal translocations. These results reveal important links between 3'ss control and ATM-dependent responses to double-strand DNA breaks, demonstrate functional plasticity of intronic variants and illustrate versatility of intronic SSOs that target pseudo-3'ss to modify gene expression.
Conflict of interest statement
A part of this work is subject to a UK patent application. The authors declare no other competing financial interests.
Figures

Similar articles
-
PUF60-activated exons uncover altered 3' splice-site selection by germline missense mutations in a single RRM.Nucleic Acids Res. 2018 Jul 6;46(12):6166-6187. doi: 10.1093/nar/gky389. Nucleic Acids Res. 2018. PMID: 29788428 Free PMC article.
-
Antisense Oligonucleotides Modulating Activation of a Nonsense-Mediated RNA Decay Switch Exon in the ATM Gene.Nucleic Acid Ther. 2016 Dec;26(6):392-400. doi: 10.1089/nat.2016.0635. Epub 2016 Sep 22. Nucleic Acid Ther. 2016. PMID: 27658045 Free PMC article.
-
Splicing Enhancers at Intron-Exon Borders Participate in Acceptor Splice Sites Recognition.Int J Mol Sci. 2020 Sep 8;21(18):6553. doi: 10.3390/ijms21186553. Int J Mol Sci. 2020. PMID: 32911621 Free PMC article.
-
Pseudoexon activation in disease by non-splice site deep intronic sequence variation - wild type pseudoexons constitute high-risk sites in the human genome.Hum Mutat. 2022 Feb;43(2):103-127. doi: 10.1002/humu.24306. Epub 2021 Dec 5. Hum Mutat. 2022. PMID: 34837434 Review.
-
ATM-Dependent Phosphorylation of All Three Members of the MRN Complex: From Sensor to Adaptor.Biomolecules. 2015 Oct 23;5(4):2877-902. doi: 10.3390/biom5042877. Biomolecules. 2015. PMID: 26512707 Free PMC article. Review.
Cited by
-
Targeting the ERG oncogene with splice-switching oligonucleotides as a novel therapeutic strategy in prostate cancer.Br J Cancer. 2020 Sep;123(6):1024-1032. doi: 10.1038/s41416-020-0951-2. Epub 2020 Jun 25. Br J Cancer. 2020. PMID: 32581342 Free PMC article.
-
A spotter's guide to SNPtic exons: The common splice variants underlying some SNP-phenotype correlations.Mol Genet Genomic Med. 2022 Jan;10(1):e1840. doi: 10.1002/mgg3.1840. Epub 2021 Oct 28. Mol Genet Genomic Med. 2022. PMID: 34708937 Free PMC article.
-
PUF60-activated exons uncover altered 3' splice-site selection by germline missense mutations in a single RRM.Nucleic Acids Res. 2018 Jul 6;46(12):6166-6187. doi: 10.1093/nar/gky389. Nucleic Acids Res. 2018. PMID: 29788428 Free PMC article.
-
Antisense Oligonucleotides Modulating Activation of a Nonsense-Mediated RNA Decay Switch Exon in the ATM Gene.Nucleic Acid Ther. 2016 Dec;26(6):392-400. doi: 10.1089/nat.2016.0635. Epub 2016 Sep 22. Nucleic Acid Ther. 2016. PMID: 27658045 Free PMC article.
-
Analysis of Pathogenic Pseudoexons Reveals Novel Mechanisms Driving Cryptic Splicing.Front Genet. 2022 Jan 24;12:806946. doi: 10.3389/fgene.2021.806946. eCollection 2021. Front Genet. 2022. PMID: 35140743 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
