

Tau-driven 26S proteasome impairment and cognitive dysfunction can be prevented early in disease by activating cAMP-PKA signaling
- PMID: 26692334
- PMCID: PMC4787271
- DOI: 10.1038/nm.4011
Tau-driven 26S proteasome impairment and cognitive dysfunction can be prevented early in disease by activating cAMP-PKA signaling
Abstract
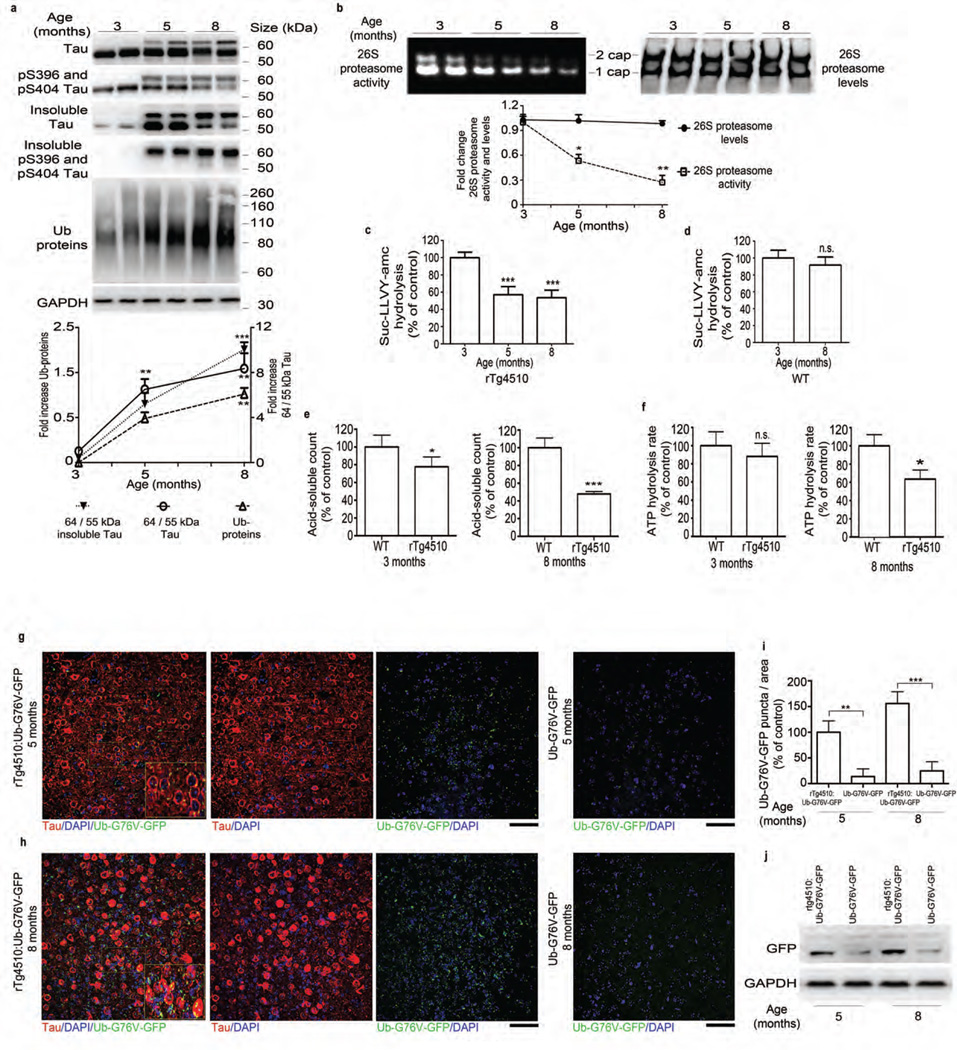
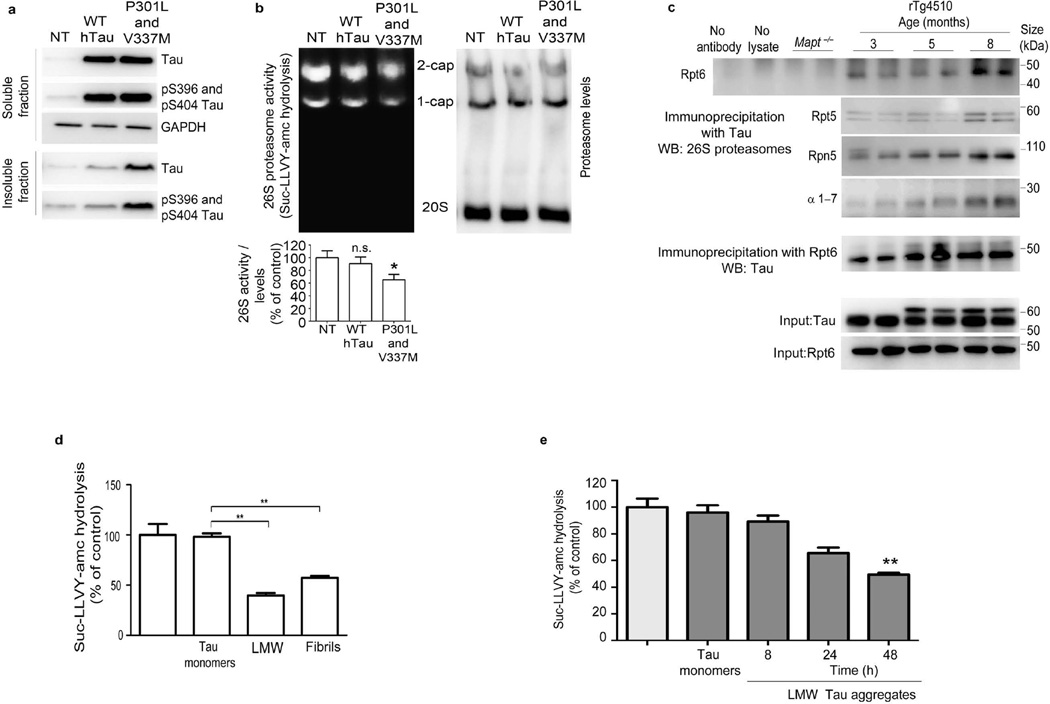
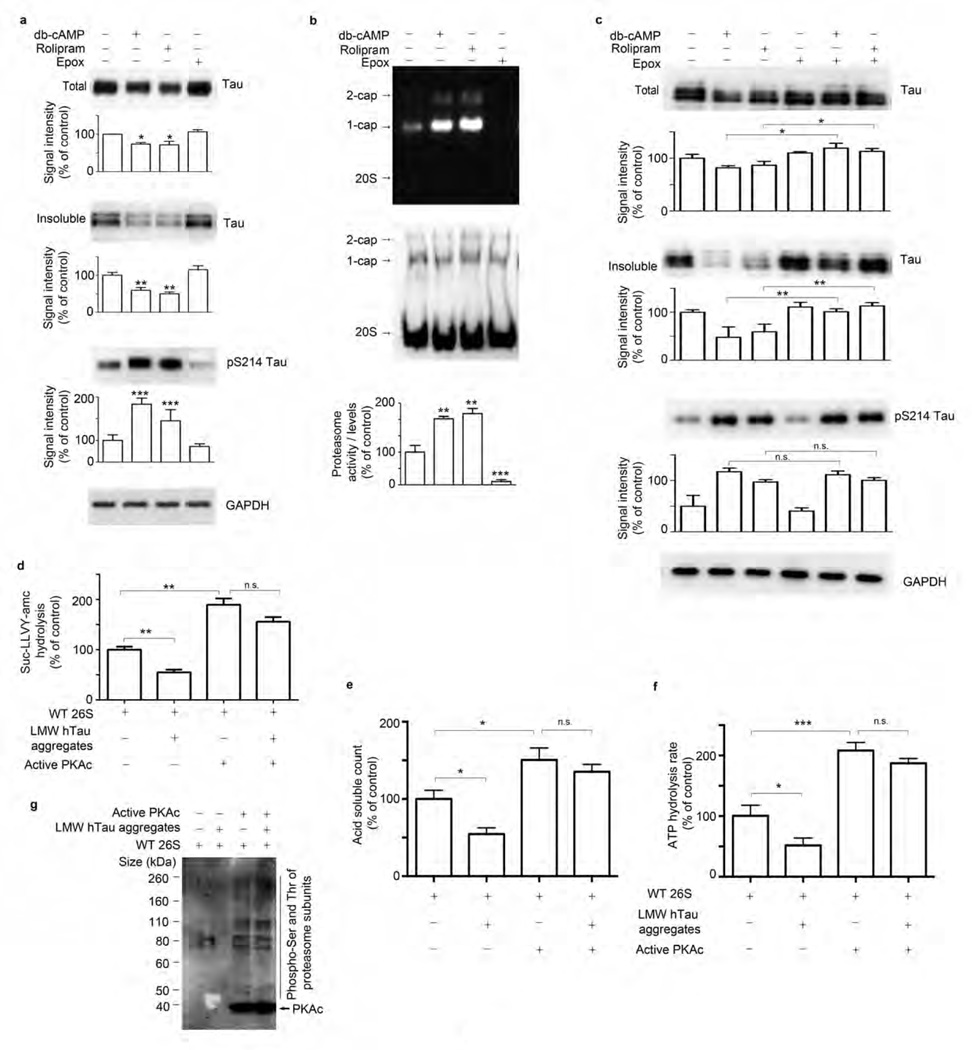
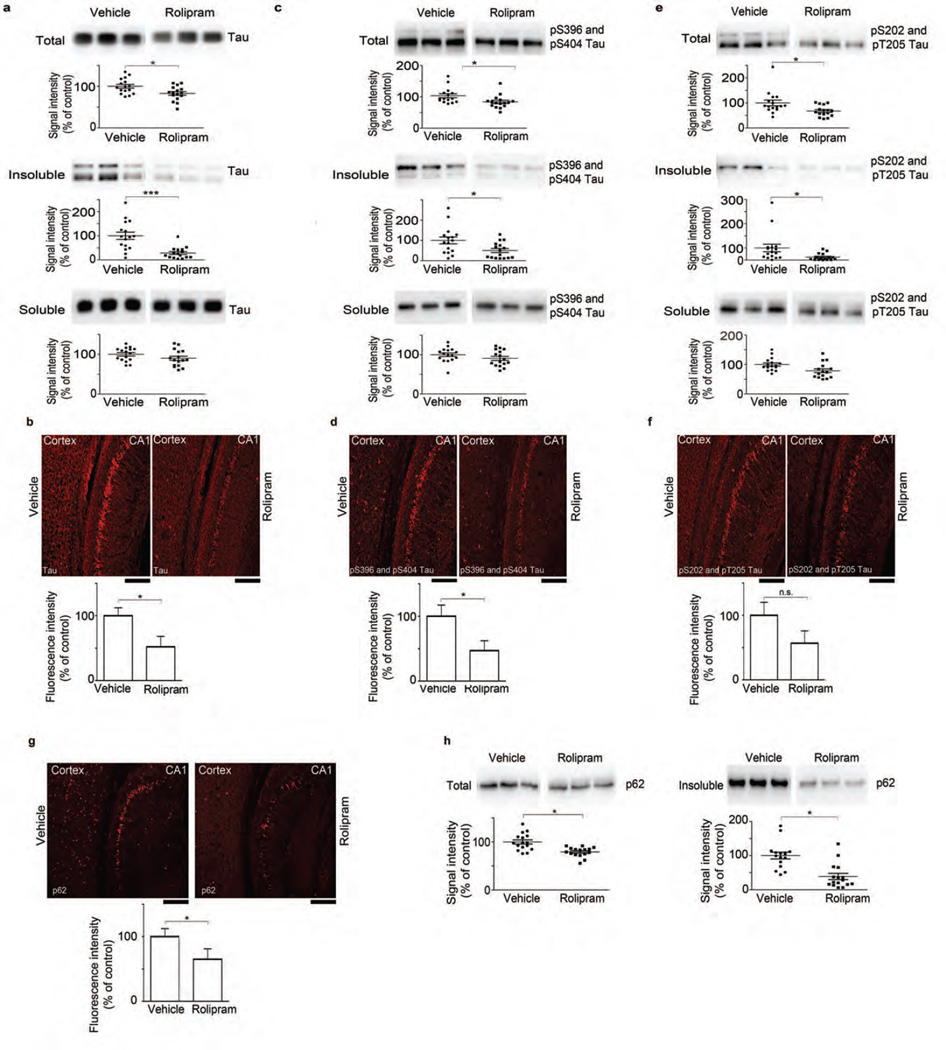
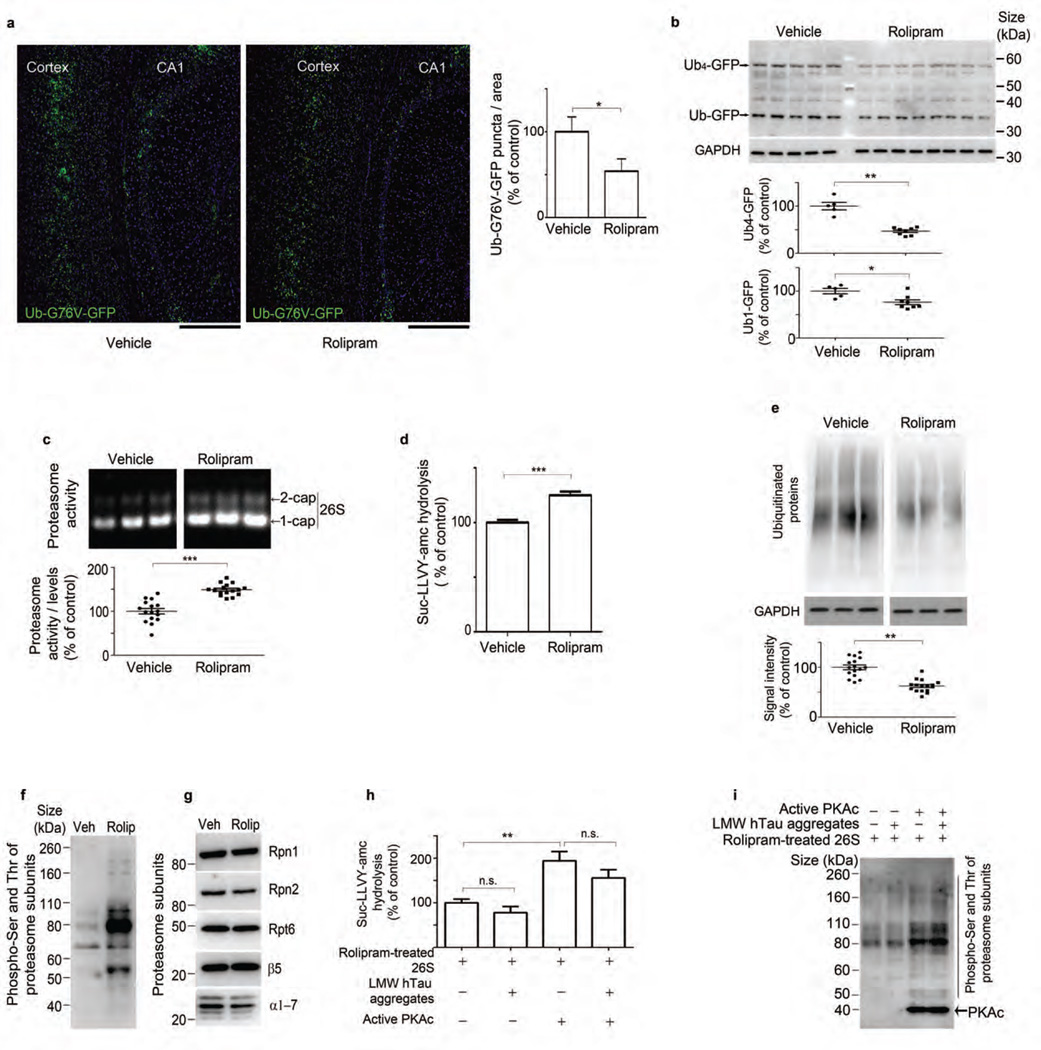
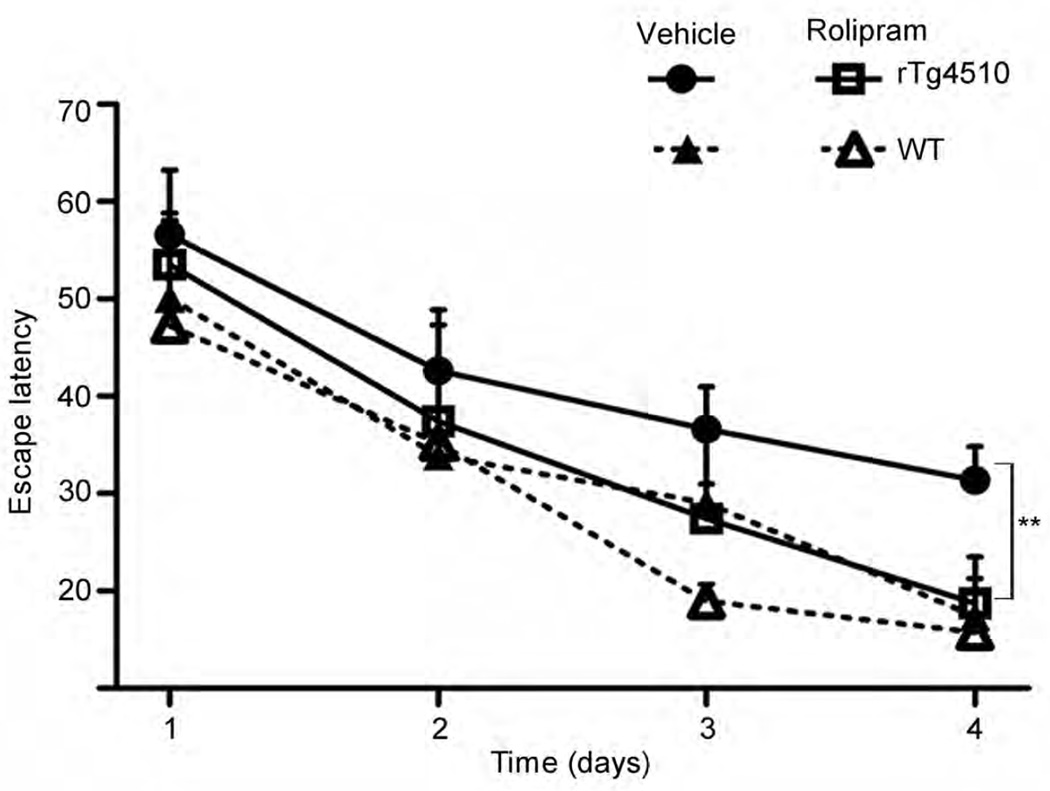
The ubiquitin proteasome system (UPS) degrades misfolded proteins including those implicated in neurodegenerative diseases. We investigated the effects of tau accumulation on proteasome function in a mouse model of tauopathy and in a cross to a UPS reporter mouse (line Ub-G76V-GFP). Accumulation of insoluble tau was associated with a decrease in the peptidase activity of brain 26S proteasomes, higher levels of ubiquitinated proteins and undegraded Ub-G76V-GFP. 26S proteasomes from mice with tauopathy were physically associated with tau and were less active in hydrolyzing ubiquitinated proteins, small peptides and ATP. 26S proteasomes from normal mice incubated with recombinant oligomers or fibrils also showed lower hydrolyzing capacity in the same assays, implicating tau as a proteotoxin. Administration of an agent that activates cAMP-protein kinase A (PKA) signaling led to attenuation of proteasome dysfunction, probably through proteasome subunit phosphorylation. In vivo, this led to lower levels of aggregated tau and improvements in cognitive performance.
Figures
Comment in
-
Tau toxicity feeds forward in frontotemporal dementia.Nat Med. 2016 Jan;22(1):24-5. doi: 10.1038/nm.4029. Nat Med. 2016. PMID: 26735407 No abstract available.
Similar articles
-
cAMP-induced phosphorylation of 26S proteasomes on Rpn6/PSMD11 enhances their activity and the degradation of misfolded proteins.Proc Natl Acad Sci U S A. 2015 Dec 29;112(52):E7176-85. doi: 10.1073/pnas.1522332112. Epub 2015 Dec 15. Proc Natl Acad Sci U S A. 2015. PMID: 26669444 Free PMC article.
-
cGMP via PKG activates 26S proteasomes and enhances degradation of proteins, including ones that cause neurodegenerative diseases.Proc Natl Acad Sci U S A. 2020 Jun 23;117(25):14220-14230. doi: 10.1073/pnas.2003277117. Epub 2020 Jun 8. Proc Natl Acad Sci U S A. 2020. PMID: 32513741 Free PMC article.
-
Tau toxicity feeds forward in frontotemporal dementia.Nat Med. 2016 Jan;22(1):24-5. doi: 10.1038/nm.4029. Nat Med. 2016. PMID: 26735407 No abstract available.
-
Regulating protein breakdown through proteasome phosphorylation.Biochem J. 2017 Sep 24;474(19):3355-3371. doi: 10.1042/BCJ20160809. Biochem J. 2017. PMID: 28947610 Free PMC article. Review.
-
Tau neurotoxicity and rescue in animal models of human Tauopathies.Curr Opin Neurobiol. 2016 Feb;36:52-8. doi: 10.1016/j.conb.2015.09.004. Epub 2015 Sep 29. Curr Opin Neurobiol. 2016. PMID: 26431808 Review.
Cited by
-
Proteasomal degradation of the intrinsically disordered protein tau at single-residue resolution.Sci Adv. 2020 Jul 22;6(30):eaba3916. doi: 10.1126/sciadv.aba3916. eCollection 2020 Jul. Sci Adv. 2020. PMID: 32832664 Free PMC article.
-
Corilagin induces human glioblastoma U251 cell apoptosis by impeding activity of (immuno)proteasome.Oncol Rep. 2021 Apr;45(4):34. doi: 10.3892/or.2021.7985. Epub 2021 Mar 2. Oncol Rep. 2021. PMID: 33649855 Free PMC article.
-
The proteasome regulator PI31 is required for protein homeostasis, synapse maintenance, and neuronal survival in mice.Proc Natl Acad Sci U S A. 2019 Dec 3;116(49):24639-24650. doi: 10.1073/pnas.1911921116. Epub 2019 Nov 21. Proc Natl Acad Sci U S A. 2019. PMID: 31754024 Free PMC article.
-
Methods to Rapidly Prepare Mammalian 26S Proteasomes for Biochemical Analysis.Methods Mol Biol. 2018;1844:277-288. doi: 10.1007/978-1-4939-8706-1_18. Methods Mol Biol. 2018. PMID: 30242716 Free PMC article.
-
AAA+ Protein-Based Technologies to Counter Neurodegenerative Disease.Biophys J. 2019 Apr 23;116(8):1380-1385. doi: 10.1016/j.bpj.2019.03.007. Epub 2019 Mar 22. Biophys J. 2019. PMID: 30952364 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
