

TGF-β/Smad3 signalling regulates the transition of bone marrow-derived macrophages into myofibroblasts during tissue fibrosis
- PMID: 26684242
- PMCID: PMC4891006
- DOI: 10.18632/oncotarget.6604
TGF-β/Smad3 signalling regulates the transition of bone marrow-derived macrophages into myofibroblasts during tissue fibrosis
Abstract
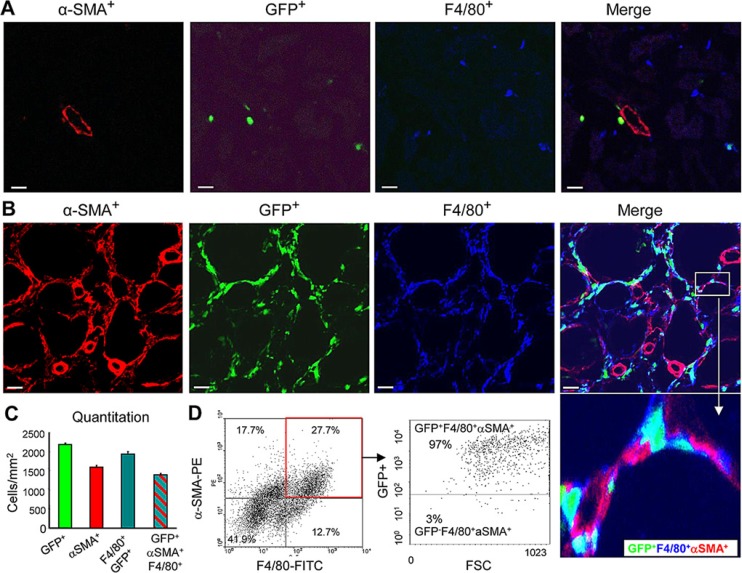
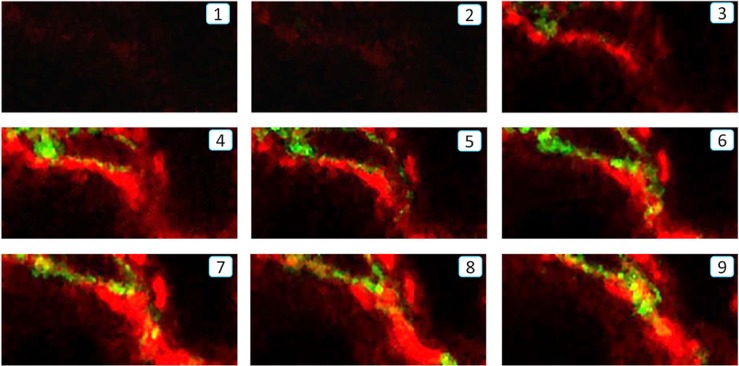
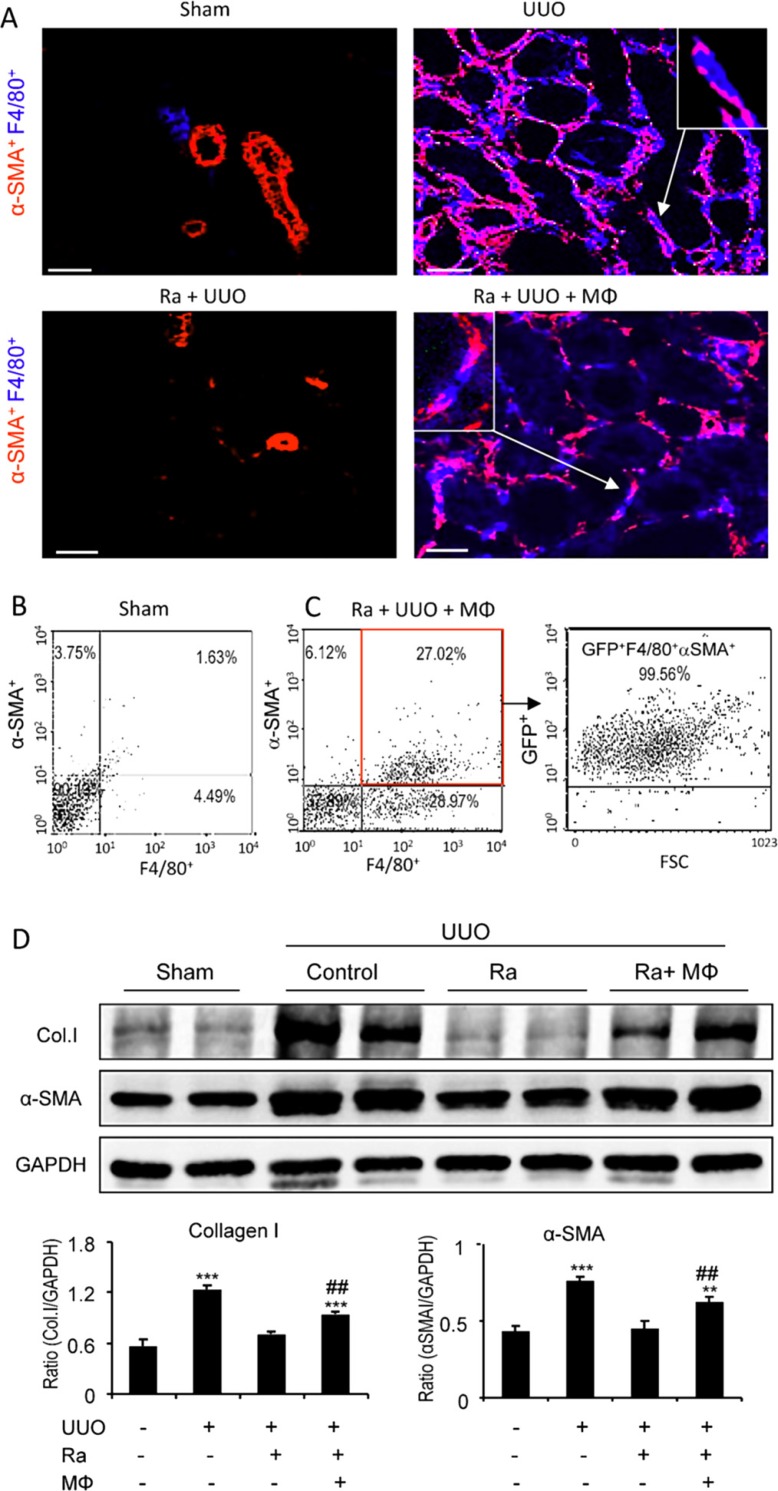
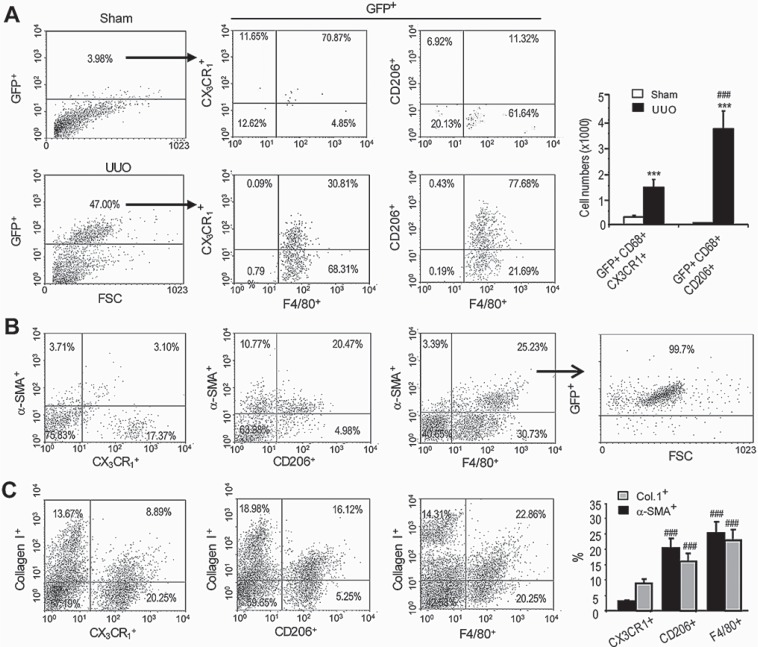
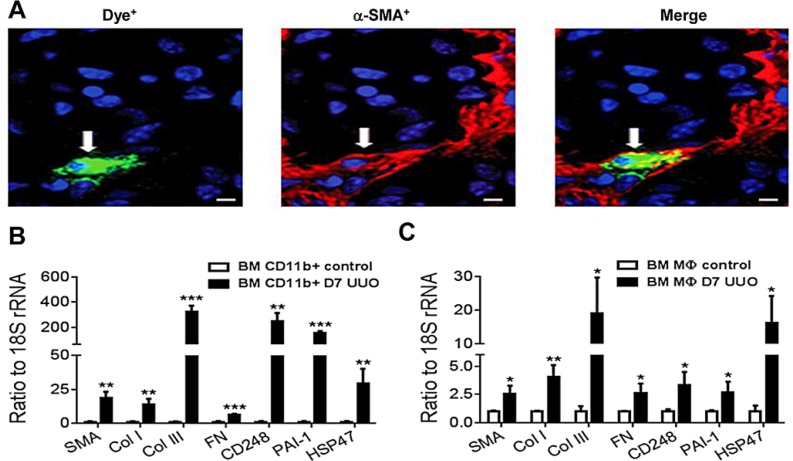
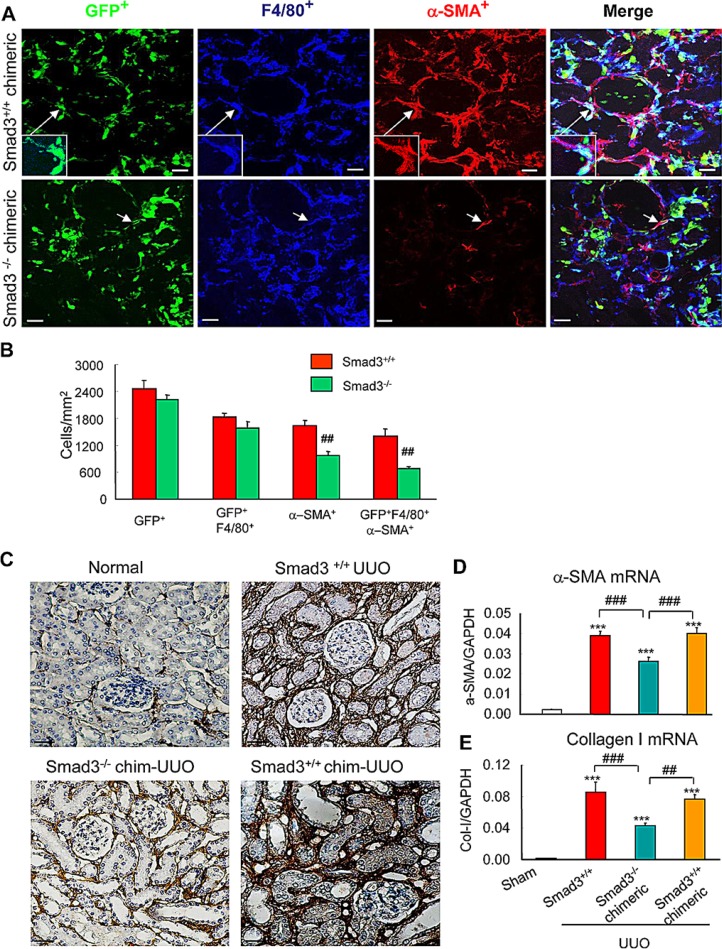
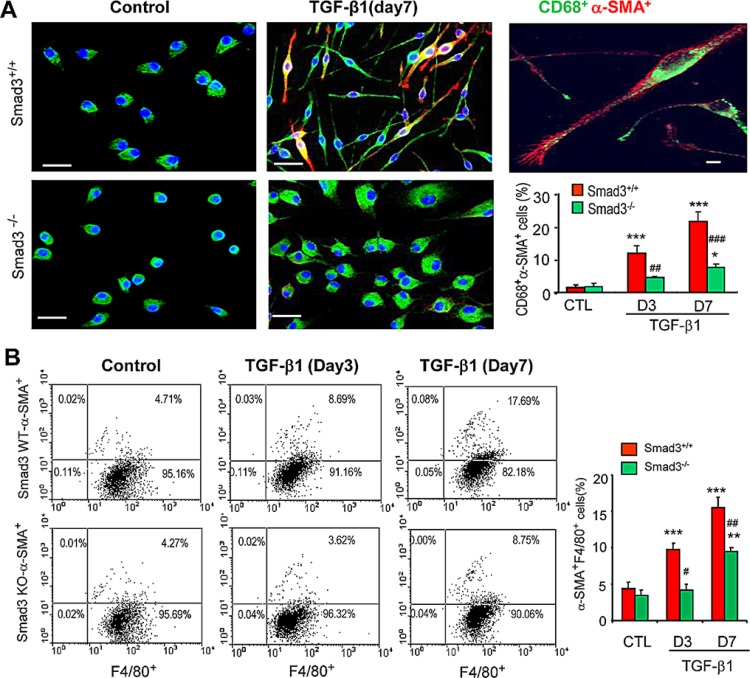
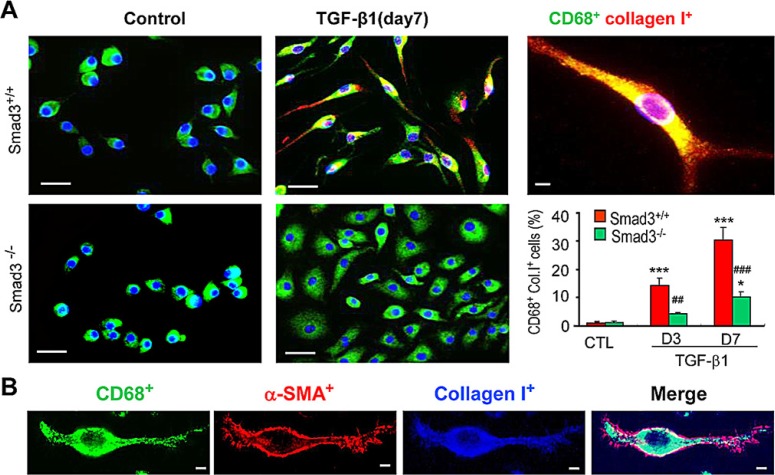
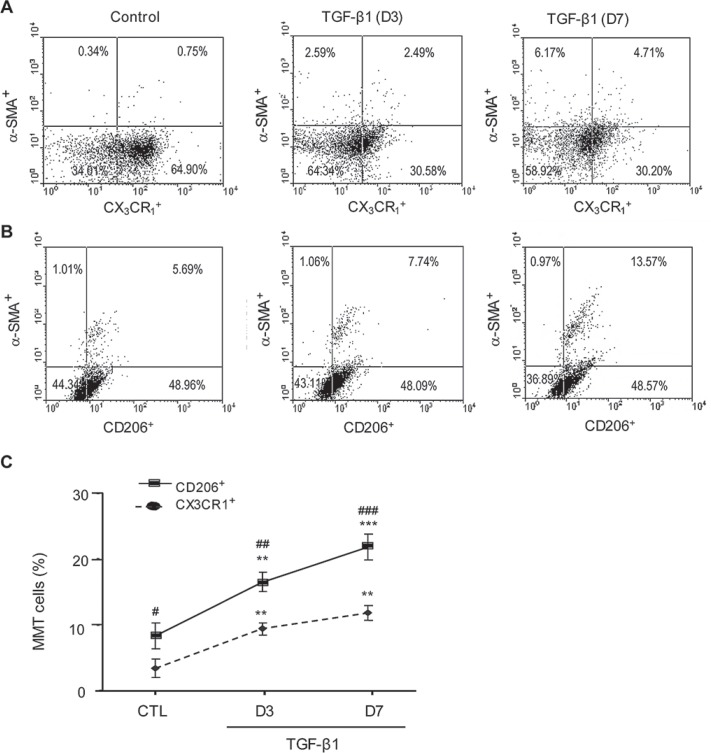
Myofibroblasts are a main cell-type of collagen-producing cells during tissue fibrosis, but their origins remains controversial. While bone marrow-derived myofibroblasts in renal fibrosis has been reported, the cell origin and mechanisms regulating their transition into myofibroblasts remain undefined. In the present study, cell lineage tracing studies by adoptive transfer of GFP+ or dye-labelled macrophages identified that monocyte/macrophages from bone marrow can give rise to myofibroblasts via the process of macrophage-myofibroblast transition (MMT) in a mouse model of unilateral ureteric obstruction. The MMT cells were a major source of collagen-producing fibroblasts in the fibrosing kidney, accounting for more than 60% of α-SMA+ myofibroblasts. The MMT process occurred predominantly within M2-type macrophages and was regulated by TGF-β/Smad3 signalling as deletion of Smad3 in the bone marrow compartment of GFP+ chimeric mice prevented the M2 macrophage transition into the MMT cells and progressive renal fibrosis. In vitro studies in Smad3 null bone marrow macrophages also showed that Smad3 was required for TGF-β1-induced MMT and collagen production. In conclusion, we have demonstrated that bone marrow-derived fibroblasts originate from the monocyte/macrophage population via a process of MMT. This process contributes to progressive renal tissue fibrosis and is regulated by TGF-β/Smad3 signalling.
Keywords: Smad3; TGF-beta; lineage tracing; macrophage-myofibroblast transition (MMT); renal fibrosis.
Conflict of interest statement
All authors declare no potential conflicts of interest.
Figures

Similar articles
-
[The effect of transforming growth factor β(1) in the transition of bone marrow-derived macrophages into myofibroblasts during renal fibrosis].Zhonghua Nei Ke Za Zhi. 2017 Aug 1;56(8):610-613. doi: 10.3760/cma.j.issn.0578-1426.2017.08.011. Zhonghua Nei Ke Za Zhi. 2017. PMID: 28789496 Chinese.
-
Inflammatory macrophages can transdifferentiate into myofibroblasts during renal fibrosis.Cell Death Dis. 2016 Dec 1;7(12):e2495. doi: 10.1038/cddis.2016.402. Cell Death Dis. 2016. PMID: 27906172 Free PMC article.
-
The proto-oncogene tyrosine protein kinase Src is essential for macrophage-myofibroblast transition during renal scarring.Kidney Int. 2018 Jan;93(1):173-187. doi: 10.1016/j.kint.2017.07.026. Epub 2017 Oct 14. Kidney Int. 2018. PMID: 29042082
-
The origin of renal fibroblasts/myofibroblasts and the signals that trigger fibrosis.Differentiation. 2016 Sep;92(3):102-107. doi: 10.1016/j.diff.2016.05.008. Epub 2016 Jun 1. Differentiation. 2016. PMID: 27262400 Review.
-
Monocyte and macrophage derived myofibroblasts: Is it fate? A review of the current evidence.Wound Repair Regen. 2021 Jul;29(4):548-562. doi: 10.1111/wrr.12946. Epub 2021 Jun 9. Wound Repair Regen. 2021. PMID: 34107123 Review.
Cited by
-
Advances in macrophage-myofibroblast transformation in fibrotic diseases.Front Immunol. 2024 Oct 9;15:1461919. doi: 10.3389/fimmu.2024.1461919. eCollection 2024. Front Immunol. 2024. PMID: 39445007 Free PMC article. Review.
-
Transforming growth factor-β in tissue fibrosis.J Exp Med. 2020 Feb 13;217(3):e20190103. doi: 10.1084/jem.20190103. Print 2020 Mar 2. J Exp Med. 2020. PMID: 32997468 Free PMC article. Review.
-
Different Roles of Resident and Non-resident Macrophages in Cardiac Fibrosis.Front Cardiovasc Med. 2022 Mar 7;9:818188. doi: 10.3389/fcvm.2022.818188. eCollection 2022. Front Cardiovasc Med. 2022. PMID: 35330948 Free PMC article. Review.
-
Deubiquitinase inhibitor PR-619 reduces Smad4 expression and suppresses renal fibrosis in mice with unilateral ureteral obstruction.PLoS One. 2018 Aug 16;13(8):e0202409. doi: 10.1371/journal.pone.0202409. eCollection 2018. PLoS One. 2018. PMID: 30114247 Free PMC article.
-
The Emerging Role of Innate Immunity in Chronic Kidney Diseases.Int J Mol Sci. 2020 Jun 4;21(11):4018. doi: 10.3390/ijms21114018. Int J Mol Sci. 2020. PMID: 32512831 Free PMC article. Review.
References
-
- Meng XM, Nikolic-Paterson DJ, Lan HY. Inflammatory processes in renal fibrosis. Nat Rev Nephrol. 2014;10:493–503. - PubMed
-
- Hewitson TD. Renal tubulointerstitial fibrosis: common but never simple. Am J Physiol Renal Physiol. 2009;296:F1239–1244. - PubMed
-
- Mack M, Yanagita M. Origin of myofibroblasts and cellular events triggering fibrosis. Kidney Int. 2015;87:297–307. - PubMed
-
- Jinde K, Nikolic-Paterson DJ, Huang XR, Sakai H, Kurokawa K, Atkins RC, Lan HY. Tubular phenotypic change in progressive tubulointerstitial fibrosis in human glomerulonephritis. Am J Kid Dis. 2001;38:761–769. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
