

miRNA-target chimeras reveal miRNA 3'-end pairing as a major determinant of Argonaute target specificity
- PMID: 26602609
- PMCID: PMC4674787
- DOI: 10.1038/ncomms9864
miRNA-target chimeras reveal miRNA 3'-end pairing as a major determinant of Argonaute target specificity
Abstract
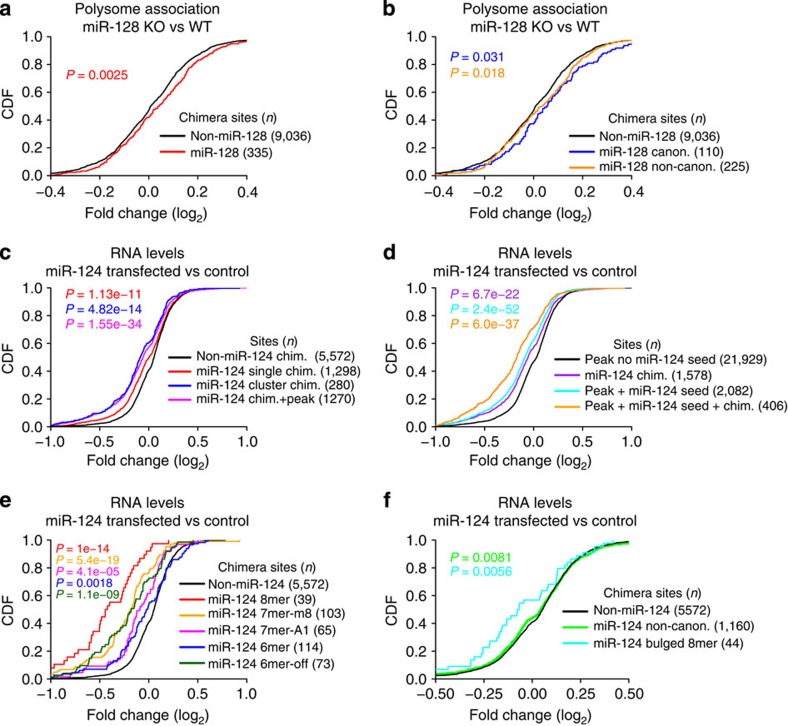
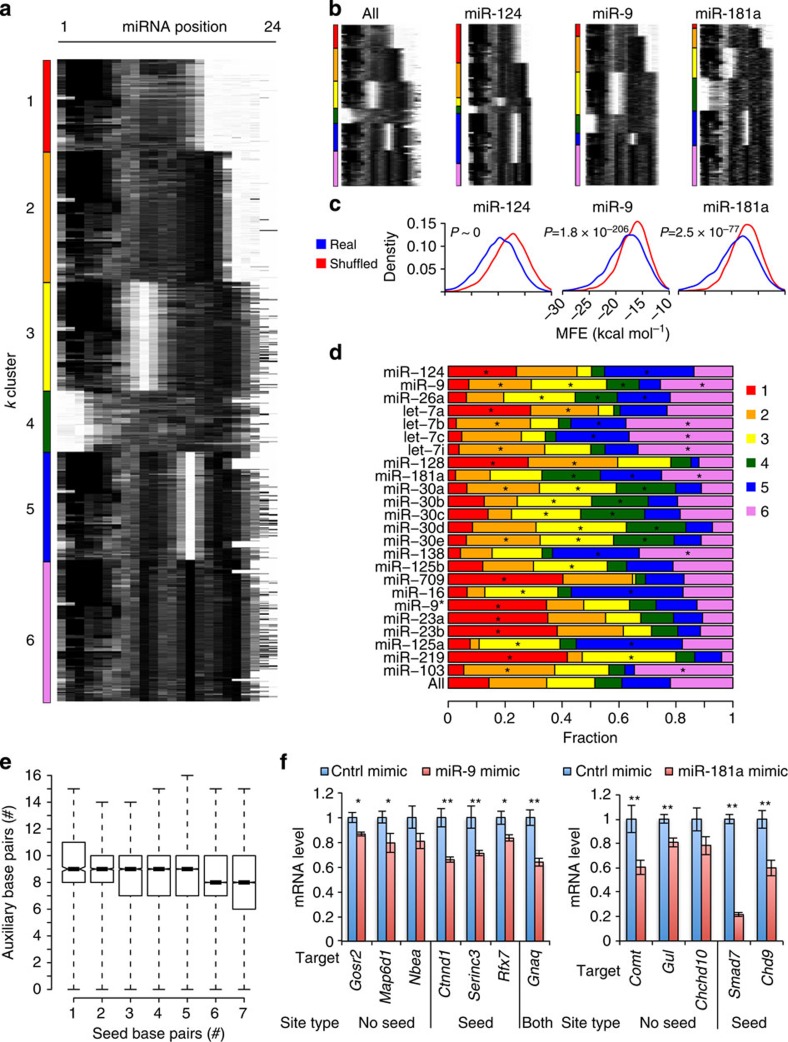
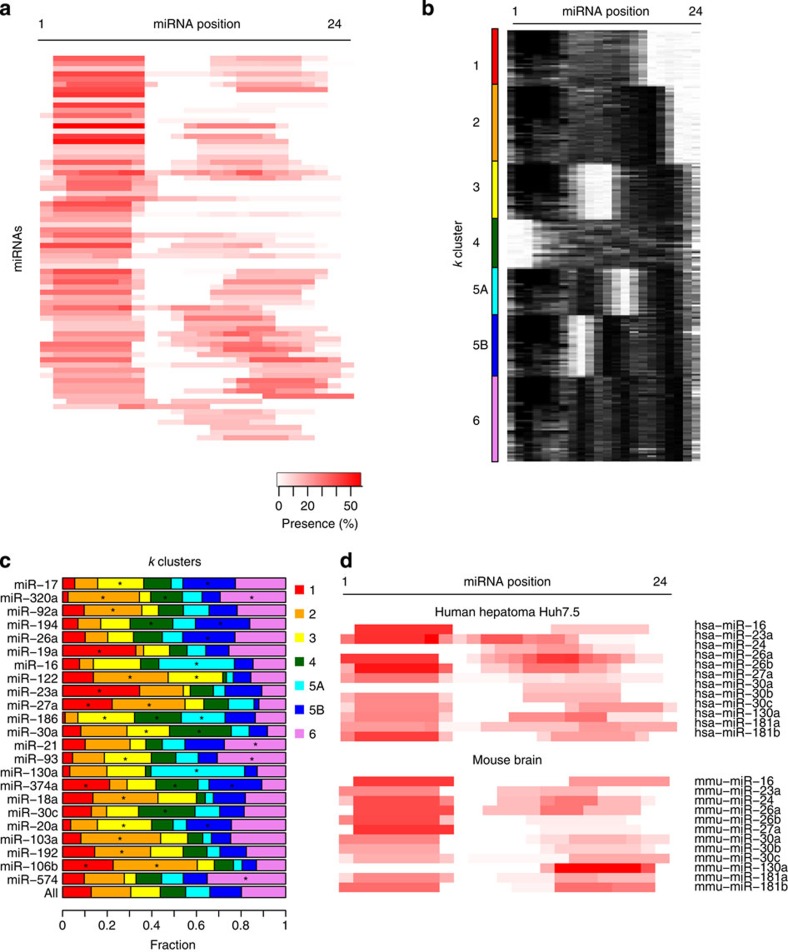
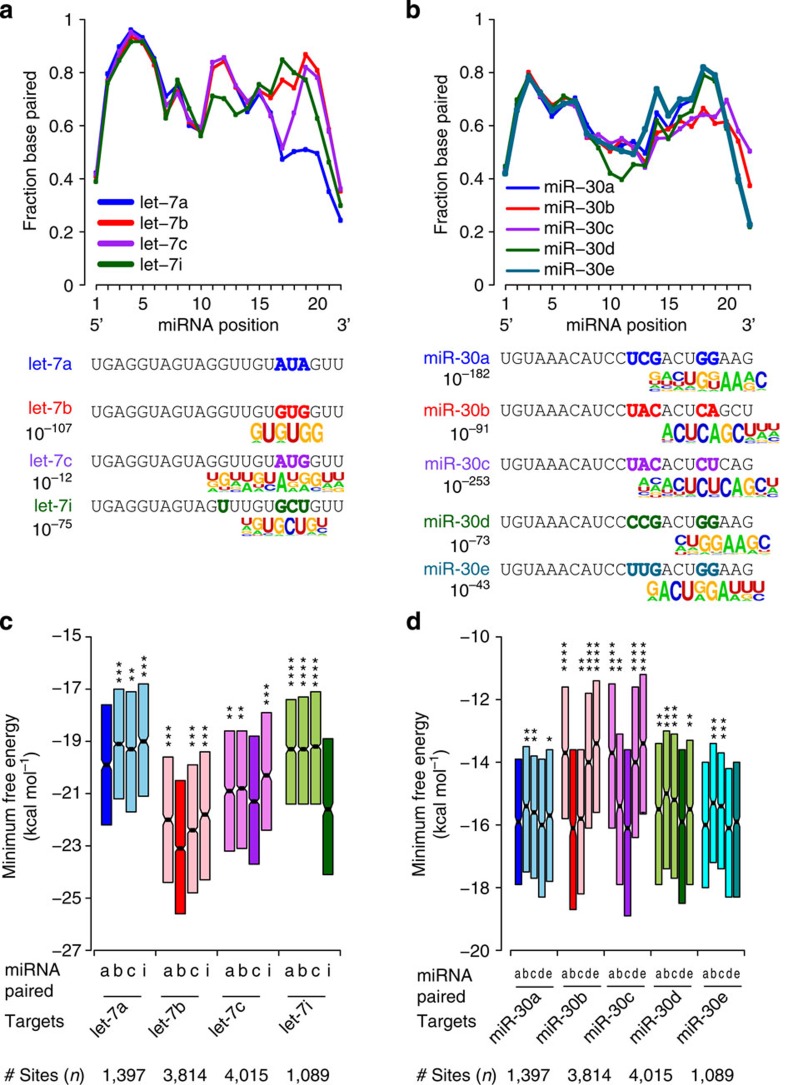
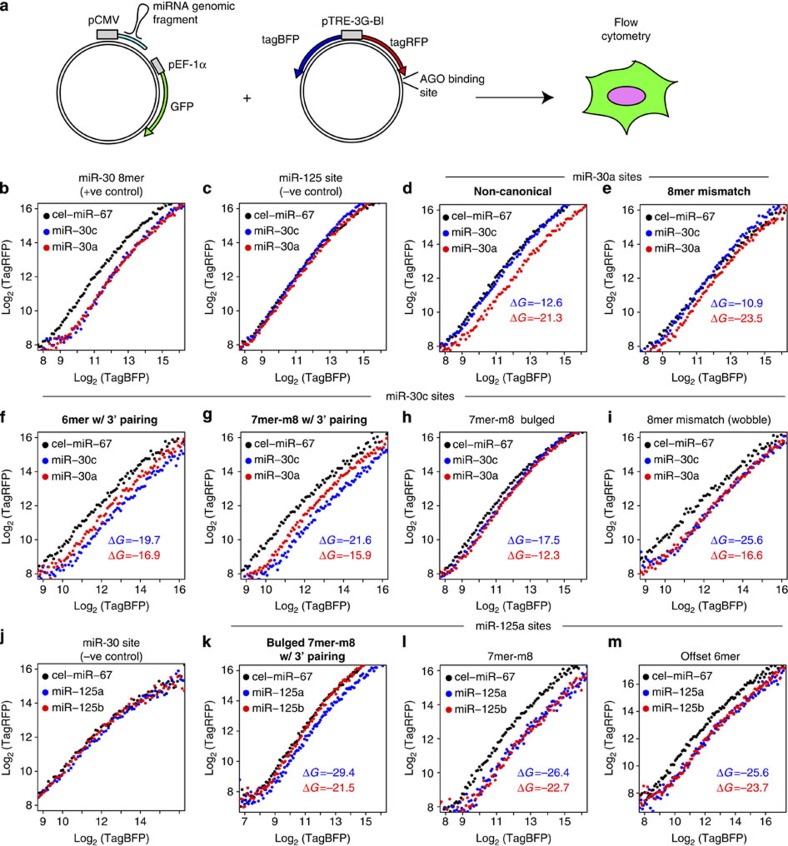
microRNAs (miRNAs) act as sequence-specific guides for Argonaute (AGO) proteins, which mediate posttranscriptional silencing of target messenger RNAs. Despite their importance in many biological processes, rules governing AGO-miRNA targeting are only partially understood. Here we report a modified AGO HITS-CLIP strategy termed CLEAR (covalent ligation of endogenous Argonaute-bound RNAs)-CLIP, which enriches miRNAs ligated to their endogenous mRNA targets. CLEAR-CLIP mapped ∼130,000 endogenous miRNA-target interactions in mouse brain and ∼40,000 in human hepatoma cells. Motif and structural analysis define expanded pairing rules for over 200 mammalian miRNAs. Most interactions combine seed-based pairing with distinct, miRNA-specific patterns of auxiliary pairing. At some regulatory sites, this specificity confers distinct silencing functions to miRNA family members with shared seed sequences but divergent 3'-ends. This work provides a means for explicit biochemical identification of miRNA sites in vivo, leading to the discovery that miRNA 3'-end pairing is a general determinant of AGO binding specificity.
Figures



Similar articles
-
Identifying miRNA Targets Using AGO-RIPseq.Methods Mol Biol. 2018;1720:131-140. doi: 10.1007/978-1-4939-7540-2_9. Methods Mol Biol. 2018. PMID: 29236255
-
Target binding triggers hierarchical phosphorylation of human Argonaute-2 to promote target release.Elife. 2022 May 31;11:e76908. doi: 10.7554/eLife.76908. Elife. 2022. PMID: 35638597 Free PMC article.
-
Unambiguous identification of miRNA:target site interactions by different types of ligation reactions.Mol Cell. 2014 Jun 19;54(6):1042-1054. doi: 10.1016/j.molcel.2014.03.049. Epub 2014 May 22. Mol Cell. 2014. PMID: 24857550 Free PMC article.
-
MicroRNA Target Recognition: Insights from Transcriptome-Wide Non-Canonical Interactions.Mol Cells. 2016 May 31;39(5):375-81. doi: 10.14348/molcells.2016.0013. Epub 2016 Apr 27. Mol Cells. 2016. PMID: 27117456 Free PMC article. Review.
-
miRNA Targeting: Growing beyond the Seed.Trends Genet. 2019 Mar;35(3):215-222. doi: 10.1016/j.tig.2018.12.005. Epub 2019 Jan 9. Trends Genet. 2019. PMID: 30638669 Free PMC article. Review.
Cited by
-
Computational Screening to Predict MicroRNA Targets in the Flavivirus 3' UTR Genome: An Approach for Antiviral Development.Int J Mol Sci. 2024 Sep 21;25(18):10135. doi: 10.3390/ijms251810135. Int J Mol Sci. 2024. PMID: 39337625 Free PMC article.
-
Network based multifactorial modelling of miRNA-target interactions.PeerJ. 2021 Mar 19;9:e11121. doi: 10.7717/peerj.11121. eCollection 2021. PeerJ. 2021. PMID: 33777541 Free PMC article.
-
Pairing beyond the Seed Supports MicroRNA Targeting Specificity.Mol Cell. 2016 Oct 20;64(2):320-333. doi: 10.1016/j.molcel.2016.09.004. Epub 2016 Oct 6. Mol Cell. 2016. PMID: 27720646 Free PMC article.
-
Modulation of miR-181 influences dopaminergic neuronal degeneration in a mouse model of Parkinson's disease.Mol Ther Nucleic Acids. 2022 Feb 12;28:1-15. doi: 10.1016/j.omtn.2022.02.007. eCollection 2022 Jun 14. Mol Ther Nucleic Acids. 2022. PMID: 35280925 Free PMC article.
-
microRNA-27b regulates hepatic lipase enzyme LIPC and reduces triglyceride degradation during hepatitis C virus infection.J Biol Chem. 2022 Jun;298(6):101983. doi: 10.1016/j.jbc.2022.101983. Epub 2022 Apr 25. J Biol Chem. 2022. PMID: 35483451 Free PMC article.
References
-
- Fabian M. R., Sonenberg N. & Filipowicz W. Regulation of mRNA translation and stability by microRNAs. Annu. Rev. Biochem. 79, 351–379 (2010). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 NS081706/NS/NINDS NIH HHS/United States
- NS034389/NS/NINDS NIH HHS/United States
- R01 AI116943/AI/NIAID NIH HHS/United States
- R01 AI091707/AI/NIAID NIH HHS/United States
- AI090055/AI/NIAID NIH HHS/United States
- R01 DK085713/DK/NIDDK NIH HHS/United States
- NS081706/NS/NINDS NIH HHS/United States
- R01 NS034389/NS/NINDS NIH HHS/United States
- CA057973/CA/NCI NIH HHS/United States
- R01 AI090055/AI/NIAID NIH HHS/United States
- R56 NS034389/NS/NINDS NIH HHS/United States
- Howard Hughes Medical Institute/United States
- DK085713/DK/NIDDK NIH HHS/United States
- AI091707/AI/NIAID NIH HHS/United States
- R01 CA057973/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
