

Control of developmentally primed erythroid genes by combinatorial co-repressor actions
- PMID: 26593974
- PMCID: PMC4673834
- DOI: 10.1038/ncomms9893
Control of developmentally primed erythroid genes by combinatorial co-repressor actions
Abstract
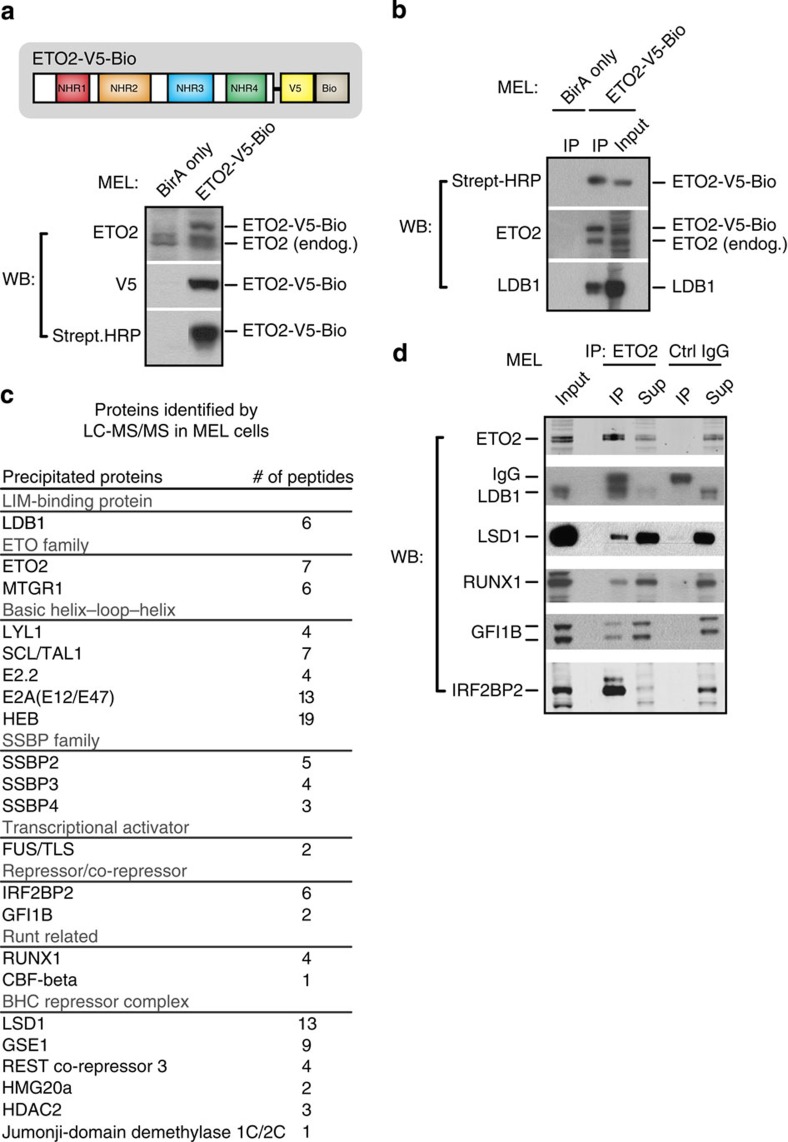
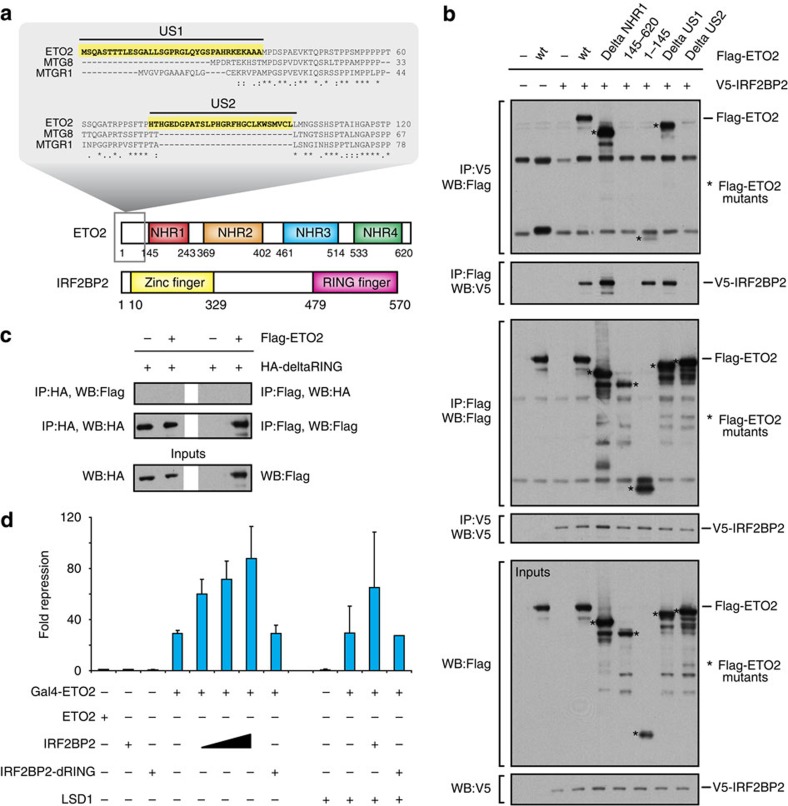
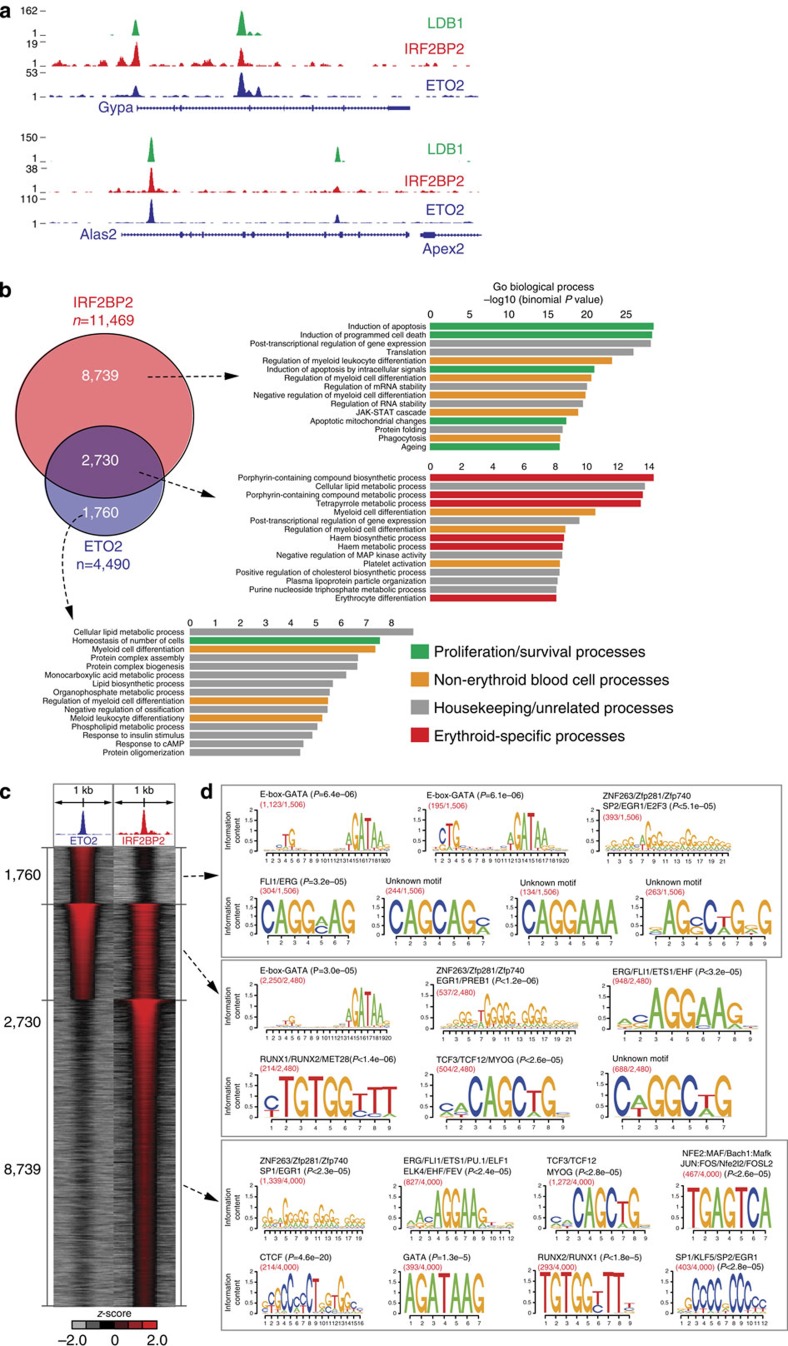
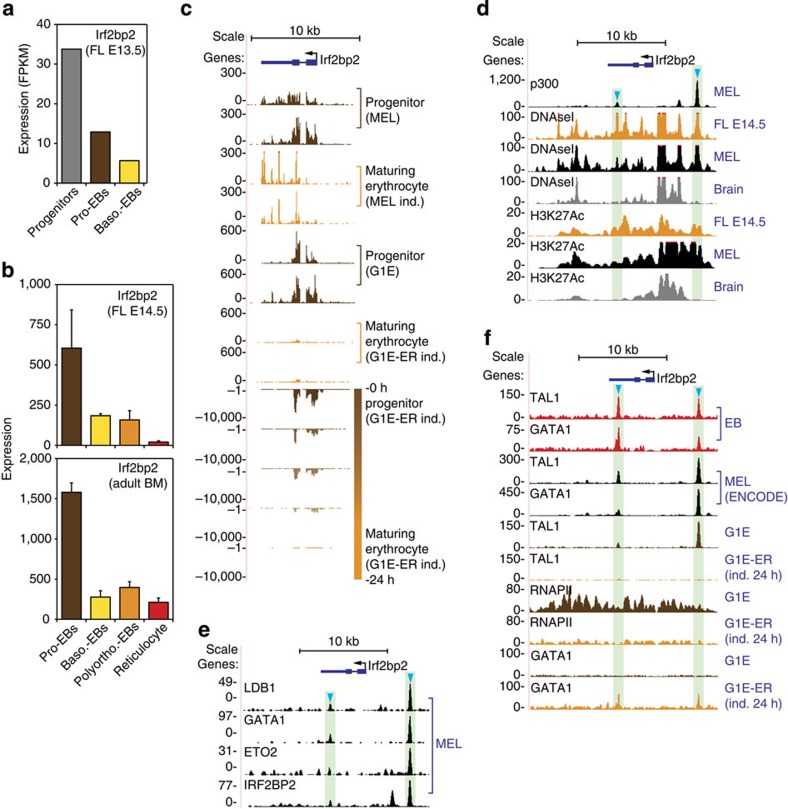
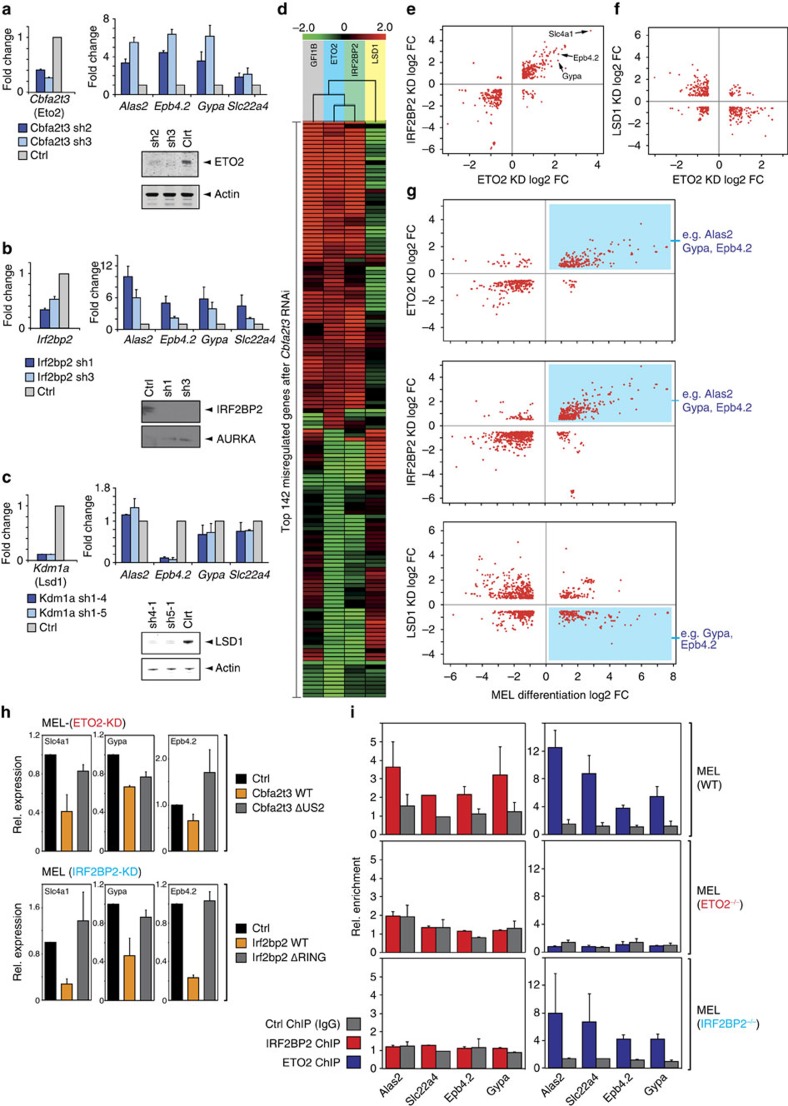
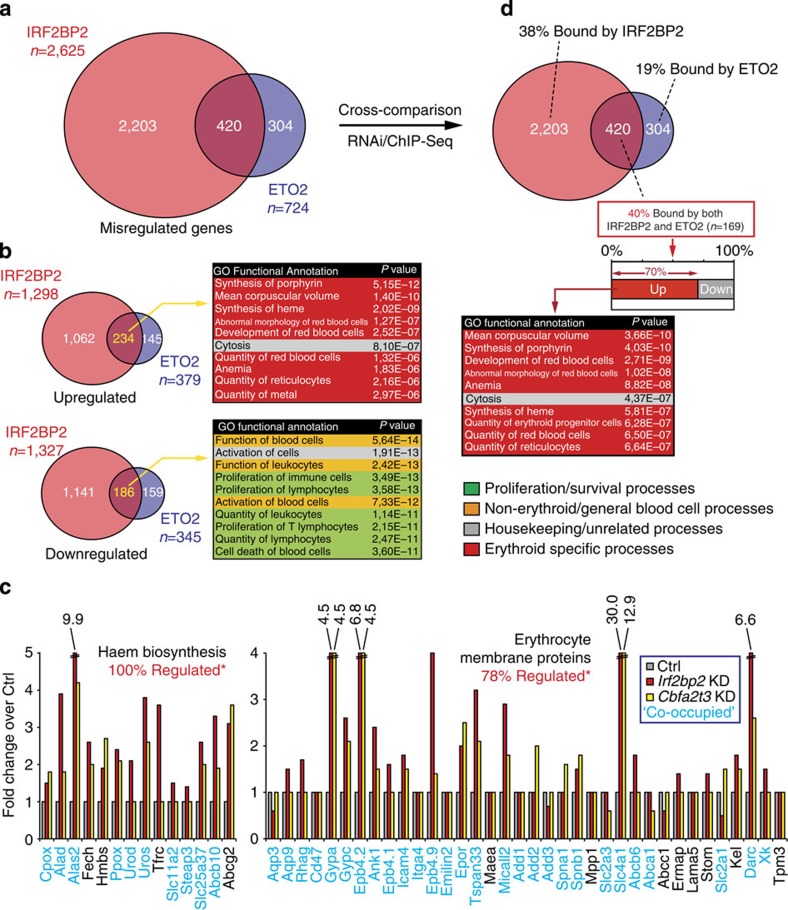
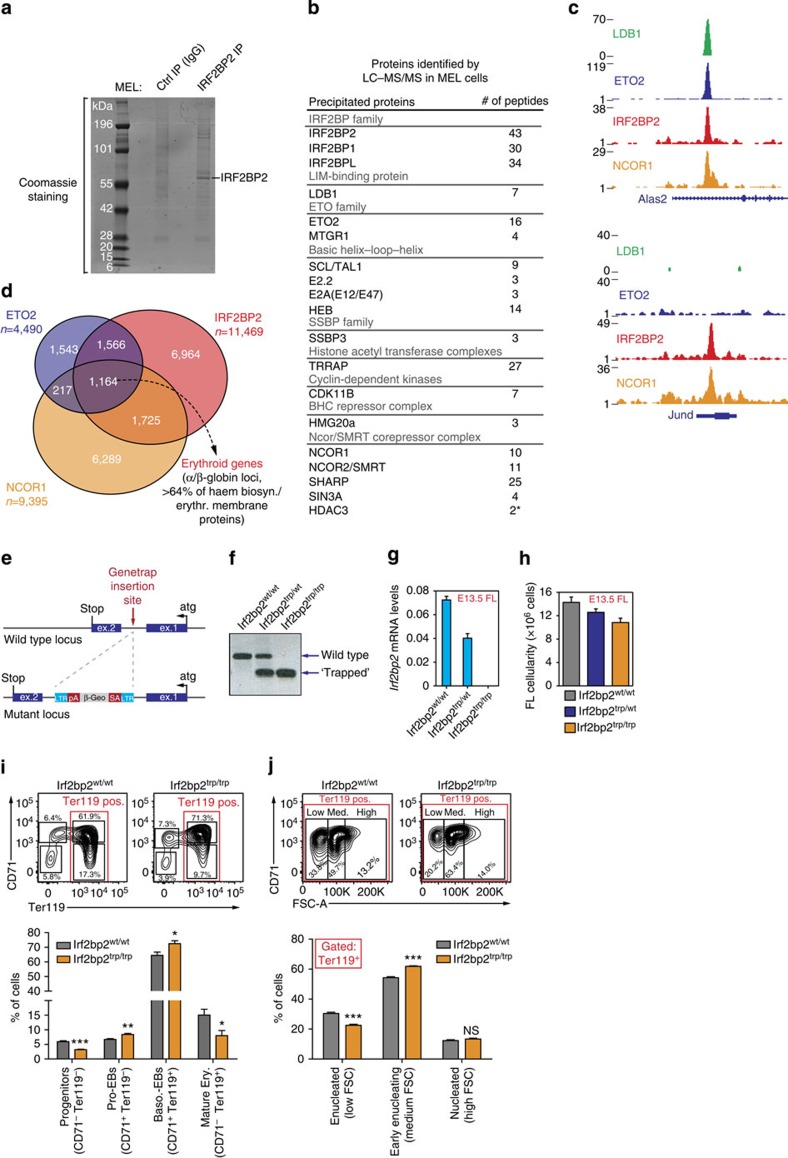
How transcription factors (TFs) cooperate within large protein complexes to allow rapid modulation of gene expression during development is still largely unknown. Here we show that the key haematopoietic LIM-domain-binding protein-1 (LDB1) TF complex contains several activator and repressor components that together maintain an erythroid-specific gene expression programme primed for rapid activation until differentiation is induced. A combination of proteomics, functional genomics and in vivo studies presented here identifies known and novel co-repressors, most notably the ETO2 and IRF2BP2 proteins, involved in maintaining this primed state. The ETO2-IRF2BP2 axis, interacting with the NCOR1/SMRT co-repressor complex, suppresses the expression of the vast majority of archetypical erythroid genes and pathways until its decommissioning at the onset of terminal erythroid differentiation. Our experiments demonstrate that multimeric regulatory complexes feature a dynamic interplay between activating and repressing components that determines lineage-specific gene expression and cellular differentiation.
Figures
Similar articles
-
Distinct Ldb1/NLI complexes orchestrate γ-globin repression and reactivation through ETO2 in human adult erythroid cells.Blood. 2011 Dec 1;118(23):6200-8. doi: 10.1182/blood-2011-06-363101. Epub 2011 Oct 18. Blood. 2011. PMID: 22010104 Free PMC article.
-
ETO-2 associates with SCL in erythroid cells and megakaryocytes and provides repressor functions in erythropoiesis.Mol Cell Biol. 2005 Dec;25(23):10235-50. doi: 10.1128/MCB.25.23.10235-10250.2005. Mol Cell Biol. 2005. PMID: 16287841 Free PMC article.
-
Embryonic erythropoiesis and hemoglobin switching require transcriptional repressor ETO2 to modulate chromatin organization.Nucleic Acids Res. 2020 Oct 9;48(18):10226-10240. doi: 10.1093/nar/gkaa736. Nucleic Acids Res. 2020. PMID: 32960220 Free PMC article.
-
Ldb1 complexes: the new master regulators of erythroid gene transcription.Trends Genet. 2014 Jan;30(1):1-9. doi: 10.1016/j.tig.2013.10.001. Epub 2013 Nov 27. Trends Genet. 2014. PMID: 24290192 Free PMC article. Review.
-
Probing the onset and regulation of erythroid cell-specific gene expression.Mt Sinai J Med. 2005 Sep;72(5):333-8. Mt Sinai J Med. 2005. PMID: 16184297 Review.
Cited by
-
IRF2BP2 prevents ox-LDL-induced inflammation and EMT in endothelial cells via regulation of KLF2.Exp Ther Med. 2021 May;21(5):481. doi: 10.3892/etm.2021.9912. Epub 2021 Mar 12. Exp Ther Med. 2021. PMID: 33767776 Free PMC article.
-
SCL/TAL1 cooperates with Polycomb RYBP-PRC1 to suppress alternative lineages in blood-fated cells.Nat Commun. 2018 Dec 18;9(1):5375. doi: 10.1038/s41467-018-07787-6. Nat Commun. 2018. PMID: 30560907 Free PMC article.
-
Interferon regulatory factor 2 binding protein 2b regulates neutrophil versus macrophage fate during zebrafish definitive myelopoiesis.Haematologica. 2020 Jan 31;105(2):325-337. doi: 10.3324/haematol.2019.217596. Print 2020. Haematologica. 2020. PMID: 31123027 Free PMC article.
-
Runx1t1 promotes the neuronal differentiation in rat hippocampus.Stem Cell Res Ther. 2020 Apr 22;11(1):160. doi: 10.1186/s13287-020-01667-x. Stem Cell Res Ther. 2020. PMID: 32321587 Free PMC article.
-
Super-enhancer-driven IRF2BP2 enhances ALK activity and promotes neuroblastoma cell proliferation.Neuro Oncol. 2024 Oct 3;26(10):1878-1894. doi: 10.1093/neuonc/noae109. Neuro Oncol. 2024. PMID: 38864832
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
