

Deletion of Methionine Sulfoxide Reductase A Does Not Affect Atherothrombosis but Promotes Neointimal Hyperplasia and Extracellular Signal-Regulated Kinase 1/2 Signaling
- PMID: 26449752
- PMCID: PMC4662651
- DOI: 10.1161/ATVBAHA.115.305857
Deletion of Methionine Sulfoxide Reductase A Does Not Affect Atherothrombosis but Promotes Neointimal Hyperplasia and Extracellular Signal-Regulated Kinase 1/2 Signaling
Abstract
Objective: Emerging evidence suggests that methionine oxidation can directly affect protein function and may be linked to cardiovascular disease. The objective of this study was to define the role of the methionine sulfoxide reductase A (MsrA) in models of vascular disease and identify its signaling pathways.
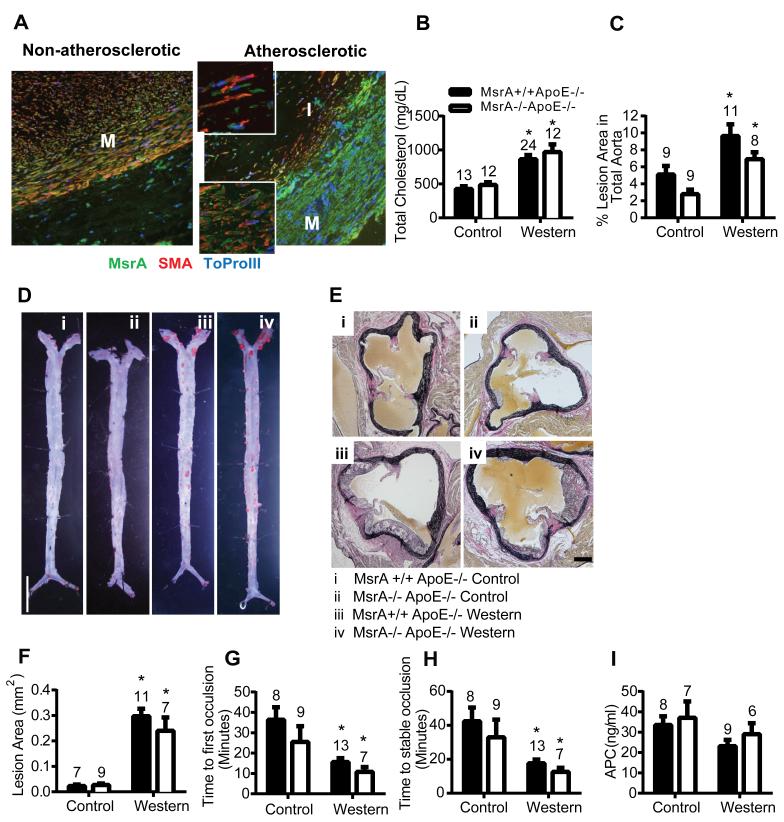
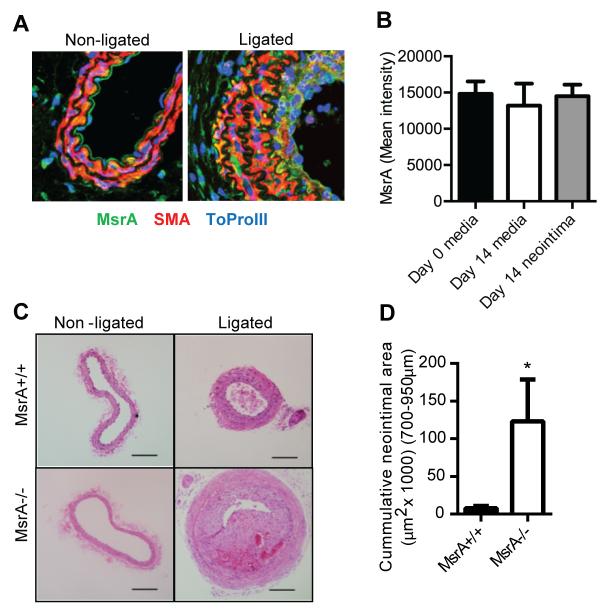
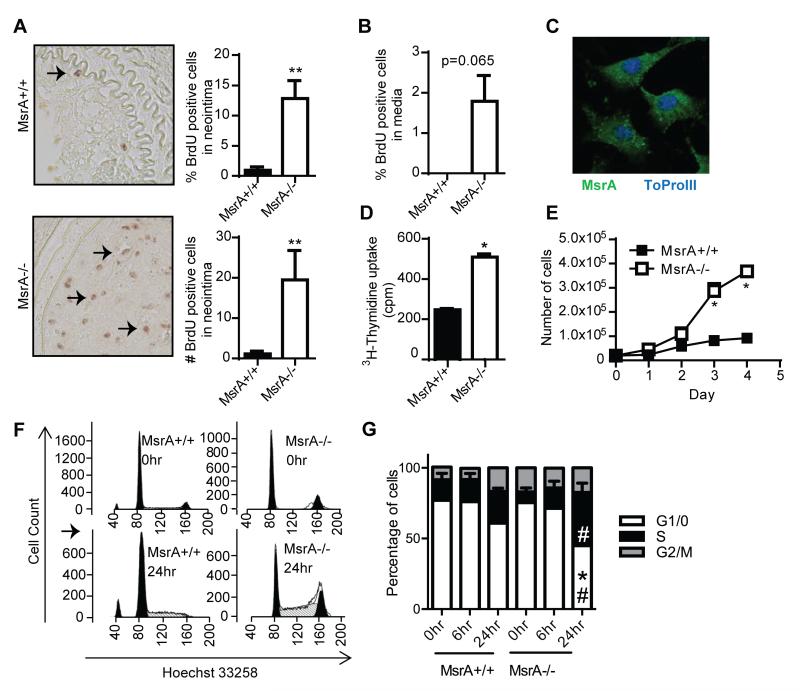
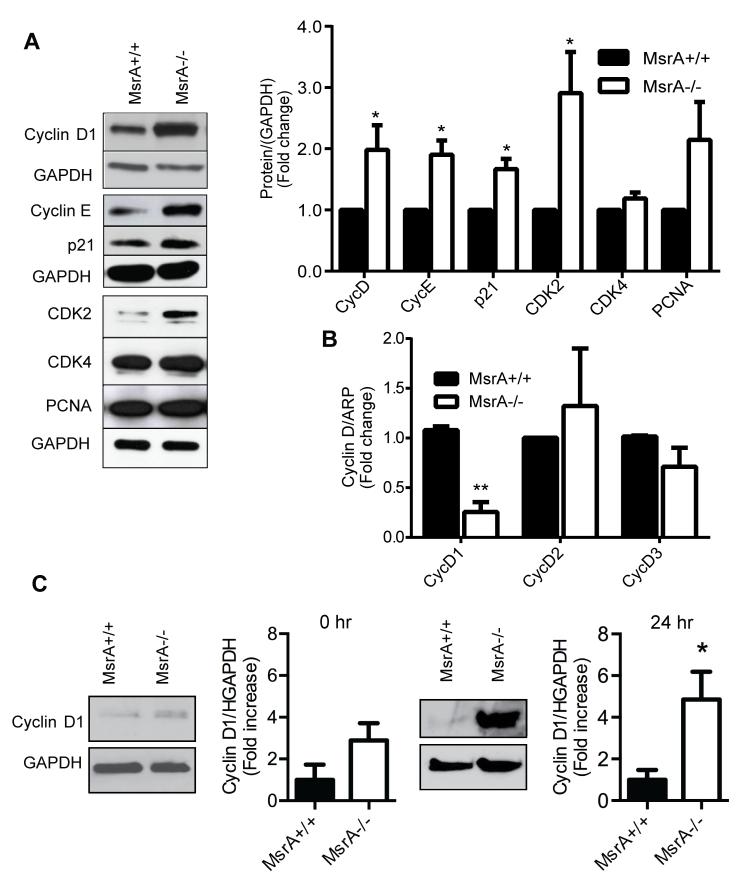
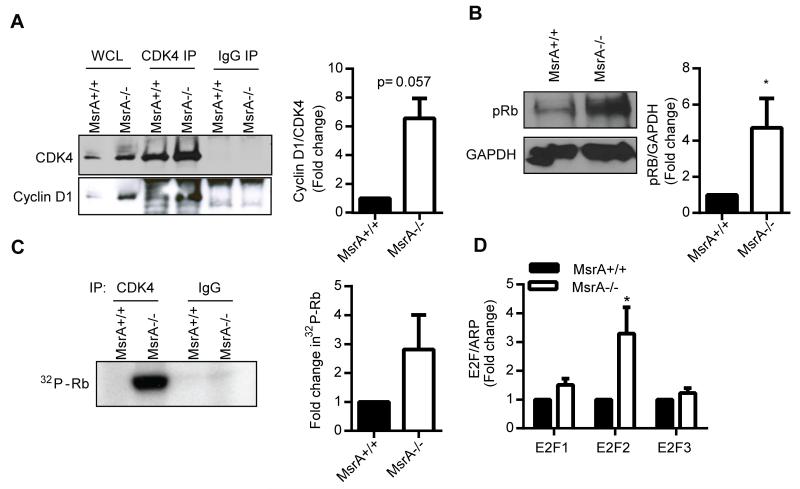
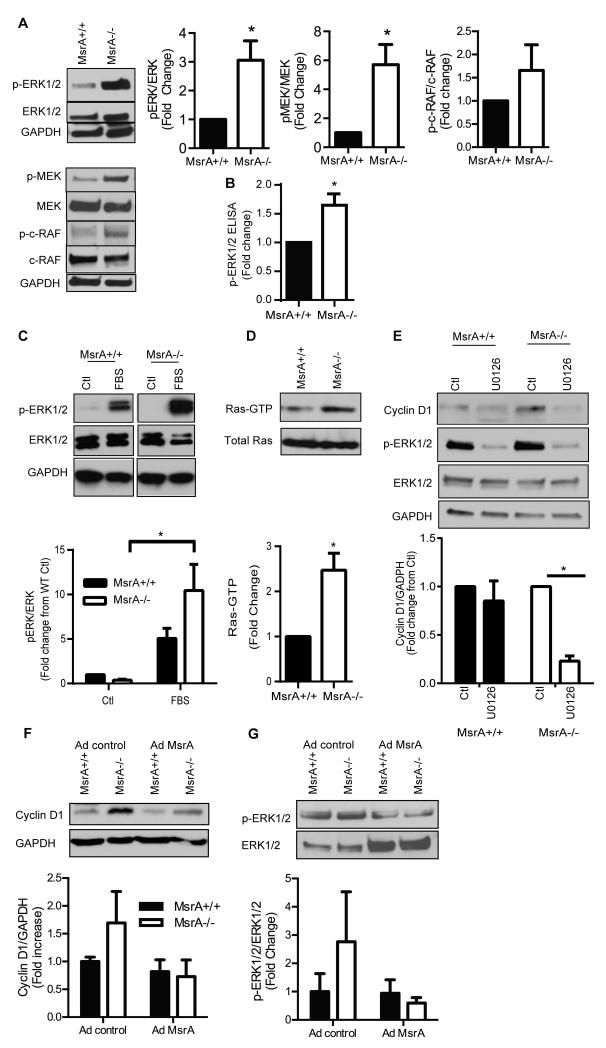
Approach and results: MsrA was readily identified in all layers of the vascular wall in human and murine arteries. Deletion of the MsrA gene did not affect atherosclerotic lesion area in apolipoprotein E-deficient mice and had no significant effect on susceptibility to experimental thrombosis after photochemical injury. In contrast, the neointimal area after vascular injury caused by complete ligation of the common carotid artery was significantly greater in MsrA-deficient than in control mice. In aortic vascular smooth muscle cells lacking MsrA, cell proliferation was significantly increased because of accelerated G1/S transition. In parallel, cyclin D1 protein and cdk4/cyclin D1 complex formation and activity were increased in MsrA-deficient vascular smooth muscle cell, leading to enhanced retinoblastoma protein phosphorylation and transcription of E2F. Finally, MsrA-deficient vascular smooth muscle cell exhibited greater activation of extracellular signal-regulated kinase 1/2 that was caused by increased activity of the Ras/Raf/mitogen-activated protein kinase signaling pathway.
Conclusions: Our findings implicate MsrA as a negative regulator of vascular smooth muscle cell proliferation and neointimal hyperplasia after vascular injury through control of the Ras/Raf/mitogen-activated protein kinase kinase/extracellular signal-regulated kinase 1/2 signaling pathway.
Keywords: ERK pathway; methionine sulfoxide reductase; neointima; oxidation-reduction; proliferation.
© 2015 American Heart Association, Inc.
Figures
Similar articles
-
Deletion of angiotensin-converting enzyme 2 promotes the development of atherosclerosis and arterial neointima formation.Cardiovasc Res. 2014 Feb 1;101(2):236-46. doi: 10.1093/cvr/cvt245. Epub 2013 Nov 4. Cardiovasc Res. 2014. PMID: 24193738
-
Blockade of the Ras-extracellular signal-regulated kinase 1/2 pathway is involved in smooth muscle 22 alpha-mediated suppression of vascular smooth muscle cell proliferation and neointima hyperplasia.Arterioscler Thromb Vasc Biol. 2010 Apr;30(4):683-91. doi: 10.1161/ATVBAHA.109.200501. Epub 2010 Feb 5. Arterioscler Thromb Vasc Biol. 2010. PMID: 20139360
-
PEA-15 (Phosphoprotein Enriched in Astrocytes 15) Is a Protective Mediator in the Vasculature and Is Regulated During Neointimal Hyperplasia.J Am Heart Assoc. 2017 Sep 11;6(9):e006936. doi: 10.1161/JAHA.117.006936. J Am Heart Assoc. 2017. PMID: 28893763 Free PMC article.
-
Involvement of heparanase in atherosclerosis and other vessel wall pathologies.Matrix Biol. 2013 Jun 24;32(5):241-51. doi: 10.1016/j.matbio.2013.03.002. Epub 2013 Mar 13. Matrix Biol. 2013. PMID: 23499530 Free PMC article. Review.
-
Notch Signaling in Endothelial Cells: Is It the Therapeutic Target for Vascular Neointimal Hyperplasia?Int J Mol Sci. 2017 Jul 25;18(8):1615. doi: 10.3390/ijms18081615. Int J Mol Sci. 2017. PMID: 28757591 Free PMC article. Review.
Cited by
-
Novel Findings From a Metabolomics Study of Left Ventricular Diastolic Function: The Bogalusa Heart Study.J Am Heart Assoc. 2020 Feb 4;9(3):e015118. doi: 10.1161/JAHA.119.015118. Epub 2020 Jan 29. J Am Heart Assoc. 2020. PMID: 31992159 Free PMC article.
-
Defective protein repair under methionine sulfoxide A deletion drives autophagy and ARE-dependent gene transcription.Redox Biol. 2018 Jun;16:401-413. doi: 10.1016/j.redox.2018.04.001. Epub 2018 Apr 3. Redox Biol. 2018. PMID: 29649787 Free PMC article.
-
Integrated analysis of the association between methionine cycle and risk of moyamoya disease.CNS Neurosci Ther. 2023 Nov;29(11):3212-3227. doi: 10.1111/cns.14254. Epub 2023 May 14. CNS Neurosci Ther. 2023. PMID: 37183324 Free PMC article. Clinical Trial.
-
Protein methionine oxidation augments reperfusion injury in acute ischemic stroke.JCI Insight. 2016;1(7):e86460. doi: 10.1172/jci.insight.86460. Epub 2016 May 19. JCI Insight. 2016. PMID: 27294204 Free PMC article.
-
Loss of methionine sulfoxide reductases increases resistance to oxidative stress.Free Radic Biol Med. 2019 Dec;145:374-384. doi: 10.1016/j.freeradbiomed.2019.10.006. Epub 2019 Oct 10. Free Radic Biol Med. 2019. PMID: 31606431 Free PMC article.
References
-
- Erickson JR, Joiner ML, Guan X, Kutschke W, Yang J, Oddis CV, Bartlett RK, Lowe JS, O'Donnell SE, Aykin-Burns N, Zimmerman MC, Zimmerman K, Ham AJ, Weiss RM, Spitz DR, Shea MA, Colbran RJ, Mohler PJ, Anderson ME. A dynamic pathway for calcium-independent activation of CaMKII by methionine oxidation. Cell. 2008;133:462–74. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
