

Detyrosinated microtubules modulate mechanotransduction in heart and skeletal muscle
- PMID: 26446751
- PMCID: PMC4633818
- DOI: 10.1038/ncomms9526
Detyrosinated microtubules modulate mechanotransduction in heart and skeletal muscle
Abstract
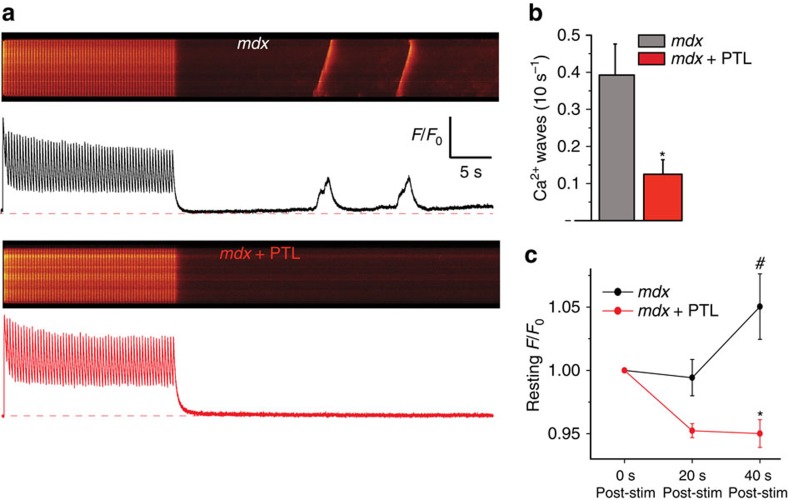
In striated muscle, X-ROS is the mechanotransduction pathway by which mechanical stress transduced by the microtubule network elicits reactive oxygen species. X-ROS tunes Ca(2+) signalling in healthy muscle, but in diseases such as Duchenne muscular dystrophy (DMD), microtubule alterations drive elevated X-ROS, disrupting Ca(2+) homeostasis and impairing function. Here we show that detyrosination, a post-translational modification of α-tubulin, influences X-ROS signalling, contraction speed and cytoskeletal mechanics. In the mdx mouse model of DMD, the pharmacological reduction of detyrosination in vitro ablates aberrant X-ROS and Ca(2+) signalling, and in vivo it protects against hallmarks of DMD, including workload-induced arrhythmias and contraction-induced injury in skeletal muscle. We conclude that detyrosinated microtubules increase cytoskeletal stiffness and mechanotransduction in striated muscle and that targeting this post-translational modification may have broad therapeutic potential in muscular dystrophies.
Conflict of interest statement
B.L.P. and C.W.W. hold a US patent for MyoTak adhesive (patent number US 20120034620 A1), licensed to IonOptix LLC. C.W.W. holds a US patent for microtubule-targeted interventions for the muscular dystrophies (patent number US 2014015664 A1). The remaining authors declare no competing financial interests.
Figures
Similar articles
-
Microtubules underlie dysfunction in duchenne muscular dystrophy.Sci Signal. 2012 Aug 7;5(236):ra56. doi: 10.1126/scisignal.2002829. Sci Signal. 2012. PMID: 22871609 Free PMC article.
-
X-ROS signaling: rapid mechano-chemo transduction in heart.Science. 2011 Sep 9;333(6048):1440-5. doi: 10.1126/science.1202768. Science. 2011. PMID: 21903813
-
Persistent upregulation of the β-tubulin tubb6, linked to muscle regeneration, is a source of microtubule disorganization in dystrophic muscle.Hum Mol Genet. 2019 Apr 1;28(7):1117-1135. doi: 10.1093/hmg/ddy418. Hum Mol Genet. 2019. PMID: 30535187 Free PMC article.
-
X-ROS signaling in the heart and skeletal muscle: stretch-dependent local ROS regulates [Ca²⁺]i.J Mol Cell Cardiol. 2013 May;58:172-81. doi: 10.1016/j.yjmcc.2012.11.011. Epub 2012 Dec 6. J Mol Cell Cardiol. 2013. PMID: 23220288 Free PMC article. Review.
-
Cytoskeletal modulation of electrical and mechanical activity in cardiac myocytes.Prog Biophys Mol Biol. 2004 Jan;84(1):29-59. doi: 10.1016/s0079-6107(03)00057-9. Prog Biophys Mol Biol. 2004. PMID: 14642867 Review.
Cited by
-
Microtubule-Mediated Misregulation of Junctophilin-2 Underlies T-Tubule Disruptions and Calcium Mishandling in mdx Mice.JACC Basic Transl Sci. 2016 Apr;1(3):122-130. doi: 10.1016/j.jacbts.2016.02.002. Epub 2016 Apr 18. JACC Basic Transl Sci. 2016. PMID: 27482548 Free PMC article.
-
Compression-dependent microtubule reinforcement enables cells to navigate confined environments.Nat Cell Biol. 2024 Sep;26(9):1520-1534. doi: 10.1038/s41556-024-01476-x. Epub 2024 Aug 19. Nat Cell Biol. 2024. PMID: 39160291
-
Mix and (mis-)match - The mechanosensing machinery in the changing environment of the developing, healthy adult and diseased heart.Biochim Biophys Acta Mol Cell Res. 2020 Mar;1867(3):118436. doi: 10.1016/j.bbamcr.2019.01.017. Epub 2019 Feb 8. Biochim Biophys Acta Mol Cell Res. 2020. PMID: 30742931 Free PMC article. Review.
-
Single-color, ratiometric biosensors for detecting signaling activities in live cells.Elife. 2018 Jul 3;7:e35458. doi: 10.7554/eLife.35458. Elife. 2018. PMID: 29968564 Free PMC article.
-
Paclitaxel-induced epithelial damage and ectopic MMP-13 expression promotes neurotoxicity in zebrafish.Proc Natl Acad Sci U S A. 2016 Apr 12;113(15):E2189-98. doi: 10.1073/pnas.1525096113. Epub 2016 Mar 28. Proc Natl Acad Sci U S A. 2016. PMID: 27035978 Free PMC article.
References
-
- Nat. Rev. Mol. Cell Biol. 10, 1–1 (2009).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
