

doi: 10.1038/nn.4085.
Modifiers of C9orf72 dipeptide repeat toxicity connect nucleocytoplasmic transport defects to FTD/ALS
Affiliations
- PMID: 26308983
- PMCID: PMC4552077
- DOI: 10.1038/nn.4085
Item in Clipboard
Modifiers of C9orf72 dipeptide repeat toxicity connect nucleocytoplasmic transport defects to FTD/ALS
Nat Neurosci.
2015 Sep.
Abstract
C9orf72 mutations are the most common cause of amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD). Dipeptide repeat proteins (DPRs) produced by unconventional translation of the C9orf72 repeat expansions cause neurodegeneration in cell culture and in animal models. We performed two unbiased screens in Saccharomyces cerevisiae and identified potent modifiers of DPR toxicity, including karyopherins and effectors of Ran-mediated nucleocytoplasmic transport, providing insight into potential disease mechanisms and therapeutic targets.
Figures
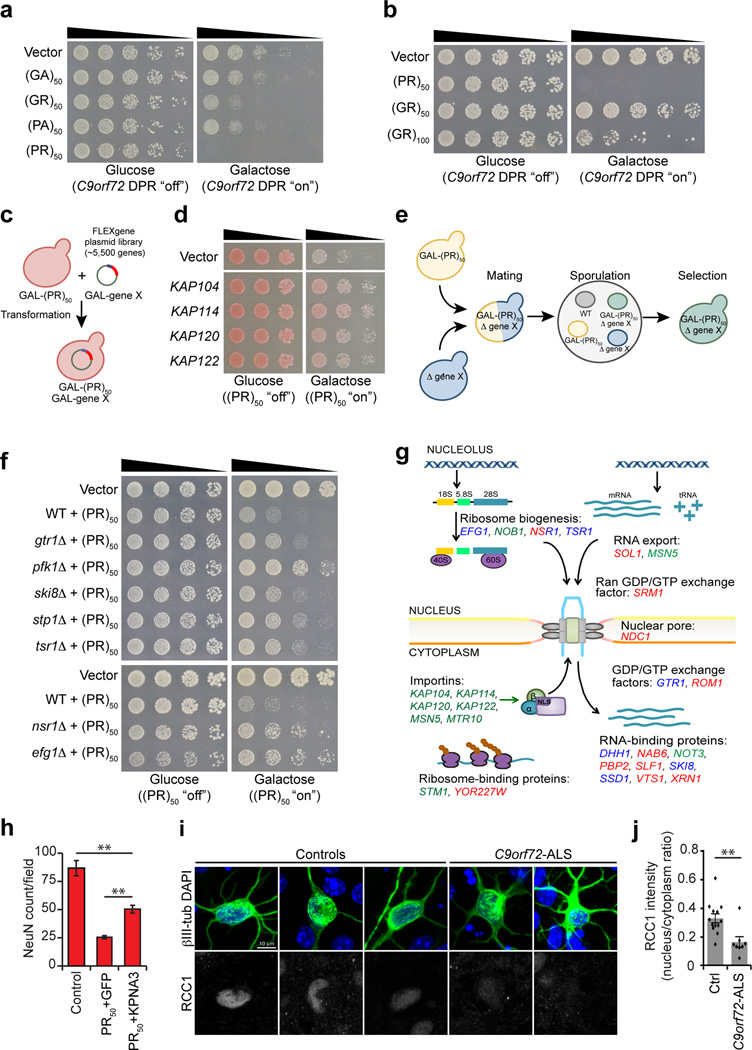
Yeast screens identify potent modifiers of C9orf72 DPR toxicity. a) Arginine-rich C9orf72 DPRs are toxic in yeast. Spotting assay demonstrates (GR)50 and (PR)50 constructs are toxic when expressed in yeast. Galactose was used to induce expression of each DPR construct and glucose was used to repress expression. Five-fold serial dilutions of yeast cells were spotted on glucose- or galactose-containing plates. b) (GR)50 is less toxic than (PR)50 and increasing the DPR length to 100 increased GR toxicity. c) Schematic of plasmid overexpression screen to identify genes that suppress or enhance (PR)50 toxicity when overexpressed. d) Examples of overexpression suppressors of (PR)50 toxicity include members of the karyopherin family of nuclear transport proteins. e) Schematic of yeast deletion screen to identify genes that suppress (PR)50 toxicity when deleted. f) Examples of deletion suppressors of (PR)50 toxicity, including gtr1Δ, a negative regulator of the Ran-GTPase cycle and nsr1Δ, a deletion of the yeast homolog of the human nucleolar protein nucleolin. g) A model depicting where the modifier genes from deletion and overexpression screens function. Genes colored blue suppressed toxicity when deleted. Genes colored red enhanced toxicity when overexpressed. Genes colored green suppressed toxicity when overexpressed. h) Upregulation of KPNA3 protects against (PR)50 toxicity in rodent neurons. Upregulation of KPNA3 more than doubled the survival of the neurons expressing (PR)50 compared to co-infection with GFP. Graph represents mean ± SEM, n = 6. ** represents p-value <0.01, by unpaired t-test. i)
C9orf72-ALS patient-derived neurons show decreased nuclear localization of RCC1 (human homolog of yeast SRM1) compared to healthy control-derived neurons. j) Quantitation of nuclear vs. cytoplasmic fluorescence intensity for RCC1. Human induced neurons from 3 healthy control subjects and 2 C9orf72-ALS patients were compared. Graph represents mean ± SEM, n = 13 (healthy controls), n = 8 (C9orf72-ALS) ** represents P <0.01, by unpaired t-test.
Comment in
-
Neurodegenerative disease: C9orf72 repeats compromise nucleocytoplasmic transport.Nat Rev Neurol. 2015 Dec;11(12):670-2. doi: 10.1038/nrneurol.2015.219. Epub 2015 Nov 3. Nat Rev Neurol. 2015. PMID: 26526532 No abstract available.
Similar articles
-
The nuclear ubiquitin ligase adaptor SPOP is a conserved regulator of C9orf72 dipeptide toxicity.Proc Natl Acad Sci U S A. 2021 Oct 5;118(40):e2104664118. doi: 10.1073/pnas.2104664118. Epub 2021 Sep 30. Proc Natl Acad Sci U S A. 2021. PMID: 34593637 Free PMC article.
-
C9orf72 ALS-FTD: recent evidence for dysregulation of the autophagy-lysosome pathway at multiple levels.Autophagy. 2021 Nov;17(11):3306-3322. doi: 10.1080/15548627.2021.1872189. Epub 2021 Feb 26. Autophagy. 2021. PMID: 33632058 Free PMC article. Review.
-
Nuclear export and translation of circular repeat-containing intronic RNA in C9ORF72-ALS/FTD.Nat Commun. 2021 Aug 13;12(1):4908. doi: 10.1038/s41467-021-25082-9. Nat Commun. 2021. PMID: 34389711 Free PMC article.
-
RAN Translation of C9orf72-Related Dipeptide Repeat Proteins in Zebrafish Recapitulates Hallmarks of Amyotrophic Lateral Sclerosis and Identifies Hypothermia as a Therapeutic Strategy.Ann Neurol. 2024 Dec;96(6):1058-1069. doi: 10.1002/ana.27068. Epub 2024 Aug 31. Ann Neurol. 2024. PMID: 39215697
-
How villains are made: The translation of dipeptide repeat proteins in C9ORF72-ALS/FTD.Gene. 2023 Mar 30;858:147167. doi: 10.1016/j.gene.2023.147167. Epub 2023 Jan 6. Gene. 2023. PMID: 36621656 Free PMC article. Review.
Cited by
-
The Development of C9orf72-Related Amyotrophic Lateral Sclerosis and Frontotemporal Dementia Disorders.Front Genet. 2020 Sep 2;11:562758. doi: 10.3389/fgene.2020.562758. eCollection 2020. Front Genet. 2020. PMID: 32983232 Free PMC article. Review.
-
Congenic expression of poly-GA but not poly-PR in mice triggers selective neuron loss and interferon responses found in C9orf72 ALS.Acta Neuropathol. 2020 Aug;140(2):121-142. doi: 10.1007/s00401-020-02176-0. Epub 2020 Jun 19. Acta Neuropathol. 2020. PMID: 32562018 Free PMC article.
-
Toxic PR Poly-Dipeptides Encoded by the C9orf72 Repeat Expansion Target LC Domain Polymers.Cell. 2016 Oct 20;167(3):789-802.e12. doi: 10.1016/j.cell.2016.10.003. Cell. 2016. PMID: 27768897 Free PMC article.
-
Poly(GR) in C9ORF72-Related ALS/FTD Compromises Mitochondrial Function and Increases Oxidative Stress and DNA Damage in iPSC-Derived Motor Neurons.Neuron. 2016 Oct 19;92(2):383-391. doi: 10.1016/j.neuron.2016.09.015. Epub 2016 Oct 6. Neuron. 2016. PMID: 27720481 Free PMC article.
-
C9orf72 dipeptides disrupt the nucleocytoplasmic transport machinery and cause TDP-43 mislocalisation to the cytoplasm.Sci Rep. 2022 Mar 21;12(1):4799. doi: 10.1038/s41598-022-08724-w. Sci Rep. 2022. PMID: 35314728 Free PMC article.
References
-
- Mori K, et al. The C9orf72 GGGGCC repeat is translated into aggregating dipeptide-repeat proteins in FTLD/ALS. Science. 2013;339:1335–1338. - PubMed
Online References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
