

Oxidative stress in β-thalassaemia and sickle cell disease
- PMID: 26285072
- PMCID: PMC4543215
- DOI: 10.1016/j.redox.2015.07.018
Oxidative stress in β-thalassaemia and sickle cell disease
Abstract
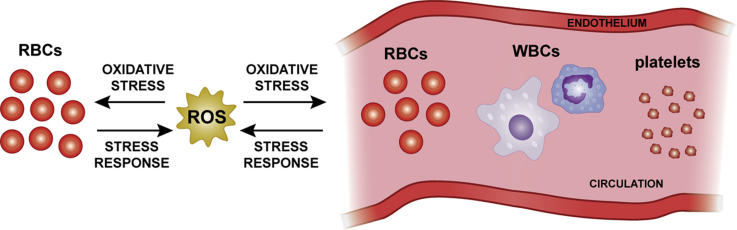
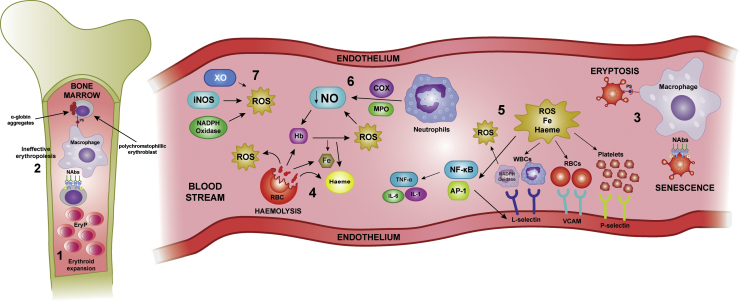
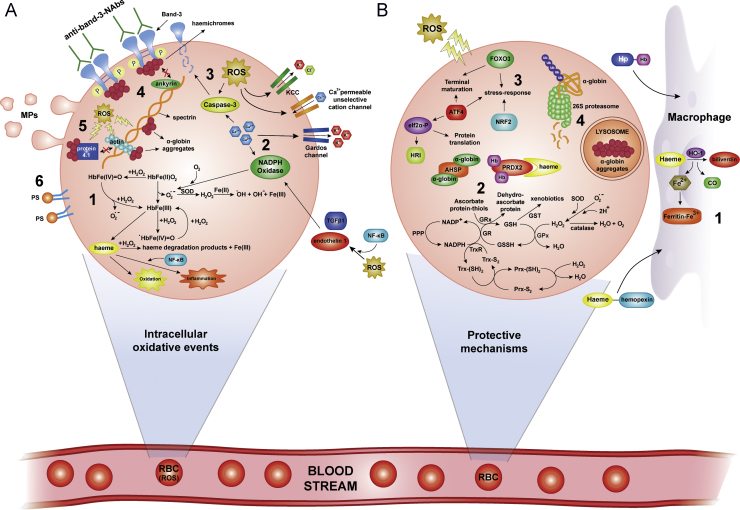
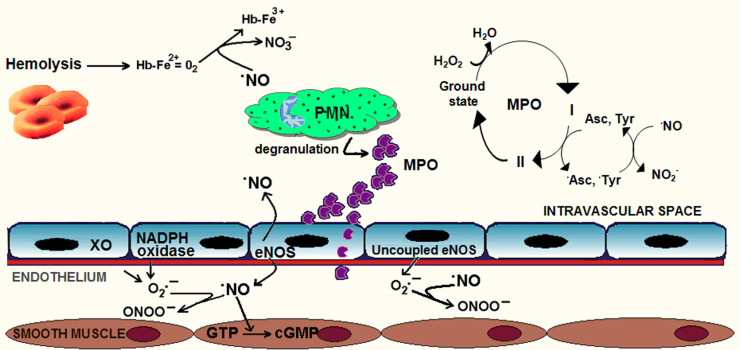
Sickle cell disease and β-thalassaemia are inherited haemoglobinopathies resulting in structural and quantitative changes in the β-globin chain. These changes lead to instability of the generated haemoglobin or to globin chain imbalance, which in turn impact the oxidative environment both intracellularly and extracellularly. The ensuing oxidative stress and the inability of the body to adequately overcome it are, to a large extent, responsible for the pathophysiology of these diseases. This article provides an overview of the main players and control mechanisms involved in the establishment of oxidative stress in these haemoglobinopathies.
Copyright © 2015 The Authors. Published by Elsevier B.V. All rights reserved.
Figures
Similar articles
-
Beta-thalassaemia and sickle cell anaemia as paradigms of hypercoagulability.Br J Haematol. 2007 Oct;139(1):3-13. doi: 10.1111/j.1365-2141.2007.06740.x. Br J Haematol. 2007. PMID: 17854302 Review.
-
Growth differentiation factor-15 in young sickle cell disease patients: relation to hemolysis, iron overload and vascular complications.Blood Cells Mol Dis. 2014 Dec;53(4):189-93. doi: 10.1016/j.bcmd.2014.07.003. Epub 2014 Jul 25. Blood Cells Mol Dis. 2014. PMID: 25065856
-
No evidence for myocardial iron overload and free iron species in multitransfused patients with sickle/beta-thalassaemia.Eur J Haematol. 2010 Jan 1;84(1):59-63. doi: 10.1111/j.1600-0609.2009.01355.x. Epub 2009 Sep 29. Eur J Haematol. 2010. PMID: 19793250
-
Erythrocytes anion transport and oxidative change in beta-thalassaemias.Cell Biol Int. 2010 May 7;34(6):655-62. doi: 10.1042/CBI20090472. Cell Biol Int. 2010. PMID: 20450494
-
[Non-transferrin-bound iron: a promising biomarker in iron overload disorders].Ned Tijdschr Geneeskd. 2013;157(49):A6258. Ned Tijdschr Geneeskd. 2013. PMID: 24299624 Review. Dutch.
Cited by
-
Oxidative Profile of Patients with Sickle Cell Disease.Med Sci (Basel). 2019 Jan 25;7(2):17. doi: 10.3390/medsci7020017. Med Sci (Basel). 2019. PMID: 30691006 Free PMC article.
-
Global analysis of erythroid cells redox status reveals the involvement of Prdx1 and Prdx2 in the severity of beta thalassemia.PLoS One. 2018 Dec 6;13(12):e0208316. doi: 10.1371/journal.pone.0208316. eCollection 2018. PLoS One. 2018. PMID: 30521599 Free PMC article.
-
Sickle Cell Trait Induces Oxidative Damage on Plasmodium falciparum Proteome at Erythrocyte Stages.Int J Mol Sci. 2019 Nov 16;20(22):5769. doi: 10.3390/ijms20225769. Int J Mol Sci. 2019. PMID: 31744112 Free PMC article.
-
Oxidative Stress in β-Thalassemia.Mol Diagn Ther. 2019 Apr;23(2):245-261. doi: 10.1007/s40291-018-0373-5. Mol Diagn Ther. 2019. PMID: 30484264 Review.
-
Mechanisms of NRF2 activation to mediate fetal hemoglobin induction and protection against oxidative stress in sickle cell disease.Exp Biol Med (Maywood). 2019 Feb;244(2):171-182. doi: 10.1177/1535370219825859. Epub 2019 Jan 23. Exp Biol Med (Maywood). 2019. PMID: 30674214 Free PMC article. Review.
References
-
- Abraham E. NF-kappaB activation. Crit. Care Med. 2000;28:N100–104. - PubMed
-
- Adamsky K., Weizer O., Amariglio N., Breda L., Harmelin A., Rivella S., Rachmilewitz E., Rechavi G. Decreased hepcidin mRNA expression in thalassemic mice. Br. J. Haematol. 2004;124:123–124. - PubMed
-
- Aidoo M., Terlouw D.J., Kolczak M.S., McElroy P.D., ter Kuile F.O., Kariuki S., Nahlen B.L., Lal A.A., Udhayakumar V. Protective effects of the sickle cell gene against malaria morbidity and mortality. Lancet. 2002;359:1311–1312. - PubMed
-
- Alam J., Killeen E., Gong P., Naquin R., Hu B., Stewart D., Ingelfinger J.R., Nath K.A. Heme activates the heme oxygenase-1 gene in renal epithelial cells by stabilizing Nrf2. Am. J. Physiol. Ren. Physiol. 2003;284:F743–752. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
