

The Shelterin TIN2 Subunit Mediates Recruitment of Telomerase to Telomeres
- PMID: 26230315
- PMCID: PMC4521702
- DOI: 10.1371/journal.pgen.1005410
The Shelterin TIN2 Subunit Mediates Recruitment of Telomerase to Telomeres
Abstract
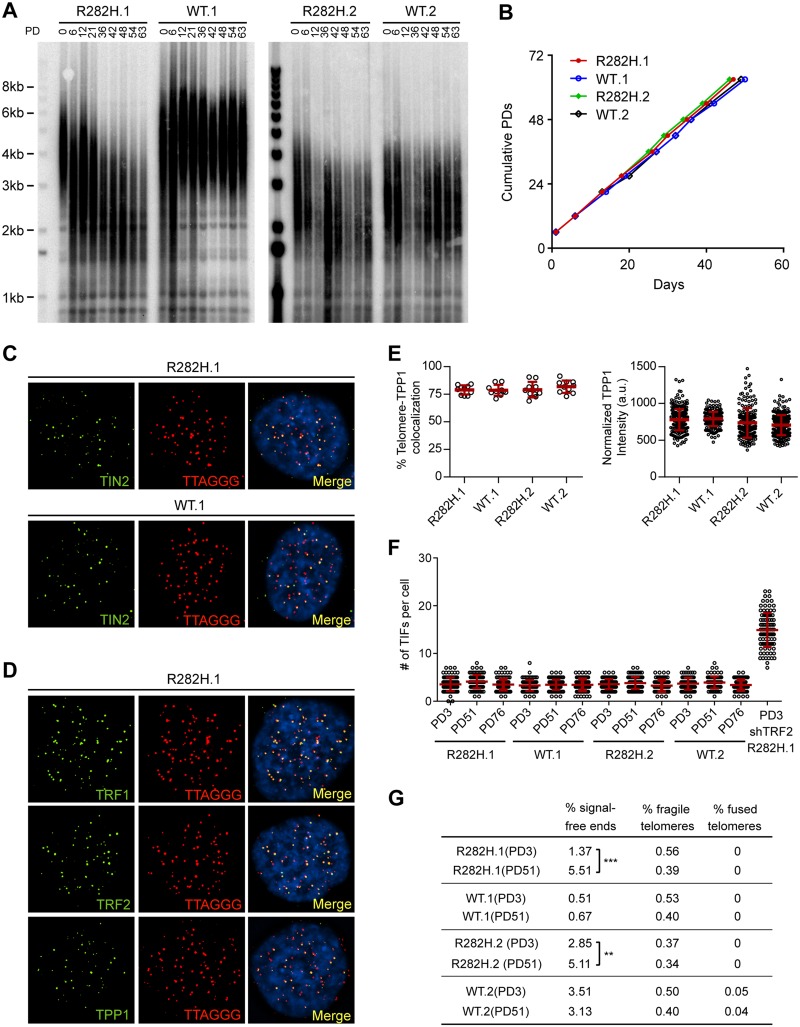
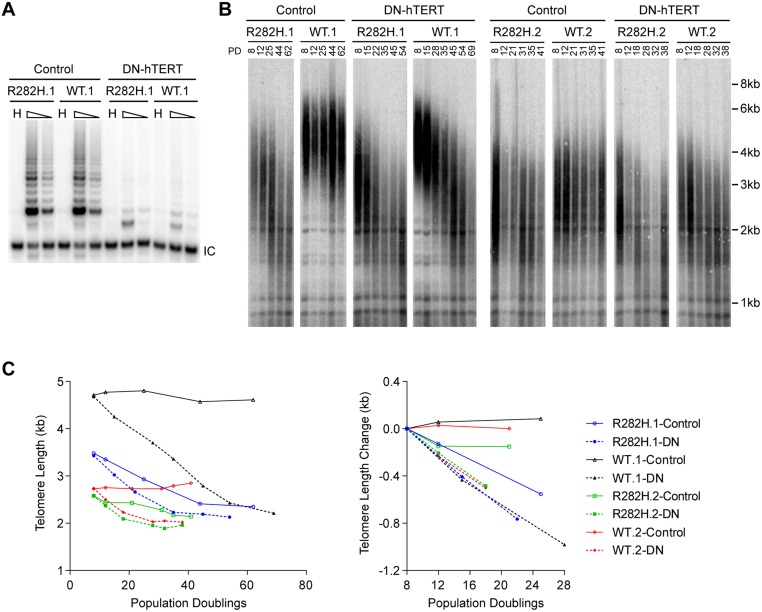
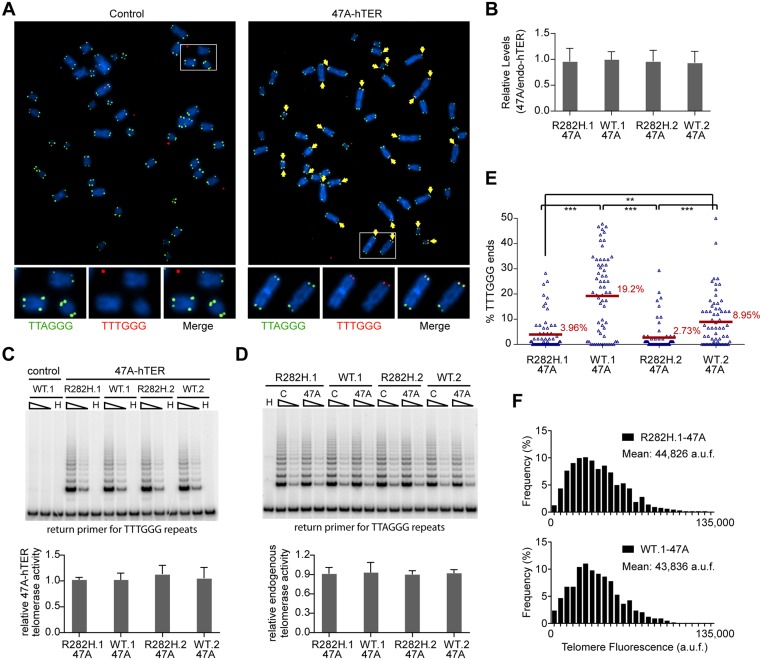
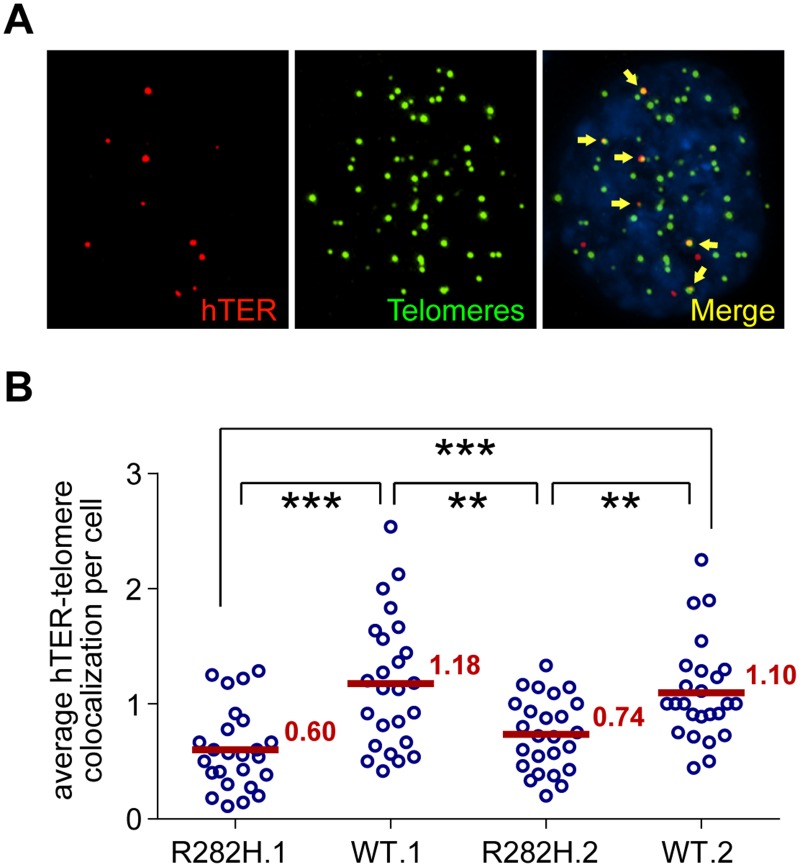
Dyskeratosis Congenita (DC) is a heritable multi-system disorder caused by abnormally short telomeres. Clinically diagnosed by the mucocutaneous symptoms, DC patients are at high risk for bone marrow failure, pulmonary fibrosis, and multiple types of cancers. We have recapitulated the most common DC-causing mutation in the shelterin component TIN2 by introducing a TIN2-R282H mutation into cultured telomerase-positive human cells via a knock-in approach. The resulting heterozygous TIN2-R282H mutation does not perturb occupancy of other shelterin components on telomeres, result in activation of telomeric DNA damage signaling or exhibit other characteristics indicative of a telomere deprotection defect. Using a novel assay that monitors the frequency and extension rate of telomerase activity at individual telomeres, we show instead that telomerase elongates telomeres at a reduced frequency in TIN2-R282H heterozygous cells; this recruitment defect is further corroborated by examining the effect of this mutation on telomerase-telomere co-localization. These observations suggest a direct role for TIN2 in mediating telomere length through telomerase, separable from its role in telomere protection.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
A TIN2 dyskeratosis congenita mutation causes telomerase-independent telomere shortening in mice.Genes Dev. 2014 Jan 15;28(2):153-66. doi: 10.1101/gad.233395.113. Genes Dev. 2014. PMID: 24449270 Free PMC article.
-
TIN2 protein dyskeratosis congenita missense mutants are defective in association with telomerase.J Biol Chem. 2011 Jul 1;286(26):23022-30. doi: 10.1074/jbc.M111.225870. Epub 2011 May 2. J Biol Chem. 2011. PMID: 21536674 Free PMC article.
-
A POT1 mutation implicates defective telomere end fill-in and telomere truncations in Coats plus.Genes Dev. 2016 Apr 1;30(7):812-26. doi: 10.1101/gad.276873.115. Epub 2016 Mar 24. Genes Dev. 2016. PMID: 27013236 Free PMC article.
-
Structural biology of telomeres and telomerase.Cell Mol Life Sci. 2020 Jan;77(1):61-79. doi: 10.1007/s00018-019-03369-x. Epub 2019 Nov 14. Cell Mol Life Sci. 2020. PMID: 31728577 Free PMC article. Review.
-
Short telomeres: from dyskeratosis congenita to sporadic aplastic anemia and malignancy.Transl Res. 2013 Dec;162(6):353-63. doi: 10.1016/j.trsl.2013.05.003. Epub 2013 Jun 1. Transl Res. 2013. PMID: 23732052 Free PMC article. Review.
Cited by
-
Role of Telomeres and Telomeric Proteins in Human Malignancies and Their Therapeutic Potential.Cancers (Basel). 2020 Jul 14;12(7):1901. doi: 10.3390/cancers12071901. Cancers (Basel). 2020. PMID: 32674474 Free PMC article. Review.
-
Telomerase Regulation from Beginning to the End.Genes (Basel). 2016 Sep 14;7(9):64. doi: 10.3390/genes7090064. Genes (Basel). 2016. PMID: 27649246 Free PMC article. Review.
-
The Role of Telomerase and Telomeres in Interstitial Lung Diseases: From Molecules to Clinical Implications.Int J Mol Sci. 2019 Jun 19;20(12):2996. doi: 10.3390/ijms20122996. Int J Mol Sci. 2019. PMID: 31248154 Free PMC article. Review.
-
Telomeres in cancer: tumour suppression and genome instability.Nat Rev Mol Cell Biol. 2017 Mar;18(3):175-186. doi: 10.1038/nrm.2016.171. Epub 2017 Jan 18. Nat Rev Mol Cell Biol. 2017. PMID: 28096526 Free PMC article. Review.
-
Fundamental mechanisms of telomerase action in yeasts and mammals: understanding telomeres and telomerase in cancer cells.Open Biol. 2017 Mar;7(3):160338. doi: 10.1098/rsob.160338. Open Biol. 2017. PMID: 28330934 Free PMC article. Review.
References
-
- Broccoli D, Smogorzewska A, Chong L, de Lange T (1997) Human telomeres contain two distinct Myb-related proteins, TRF1 and TRF2. Nat Genet 17: 231–235. - PubMed
-
- Baumann P, Cech TR (2001) Pot1, the putative telomere end-binding protein in fission yeast and humans. Science 292: 1171–1175. - PubMed
-
- Wang F, Podell ER, Zaug AJ, Yang Y, Baciu P, et al. (2007) The POT1-TPP1 telomere complex is a telomerase processivity factor. Nature 445: 506–510. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
