

An Overview of Genome Organization and How We Got There: from FISH to Hi-C
- PMID: 26223848
- PMCID: PMC4517094
- DOI: 10.1128/MMBR.00006-15
An Overview of Genome Organization and How We Got There: from FISH to Hi-C
Abstract
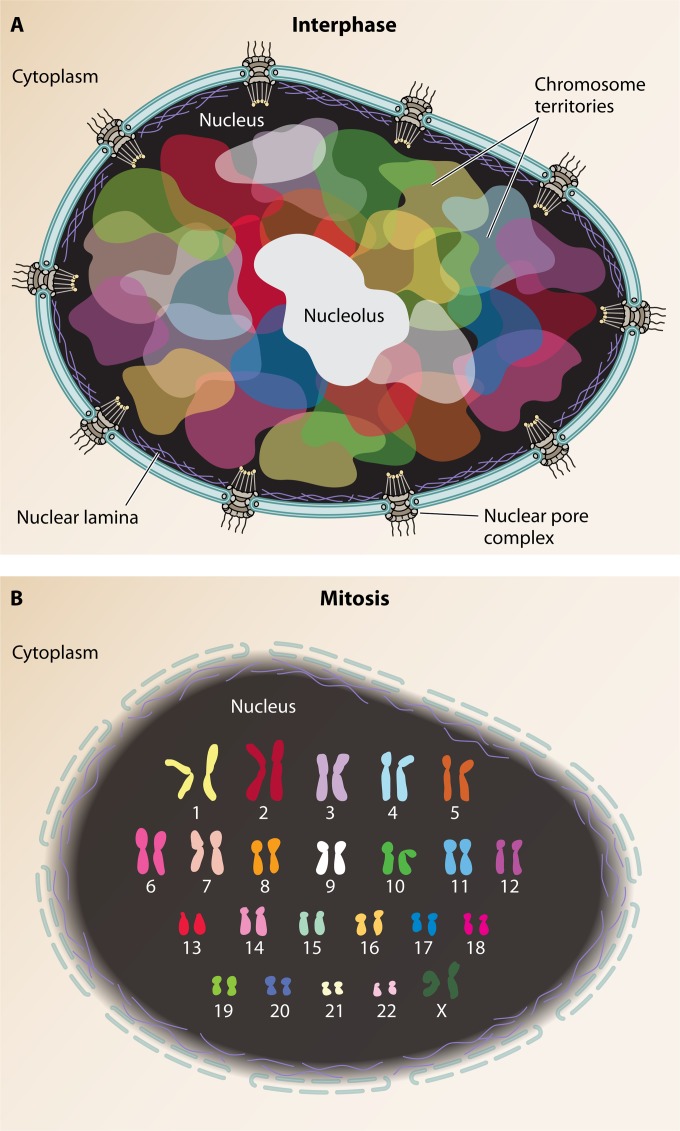
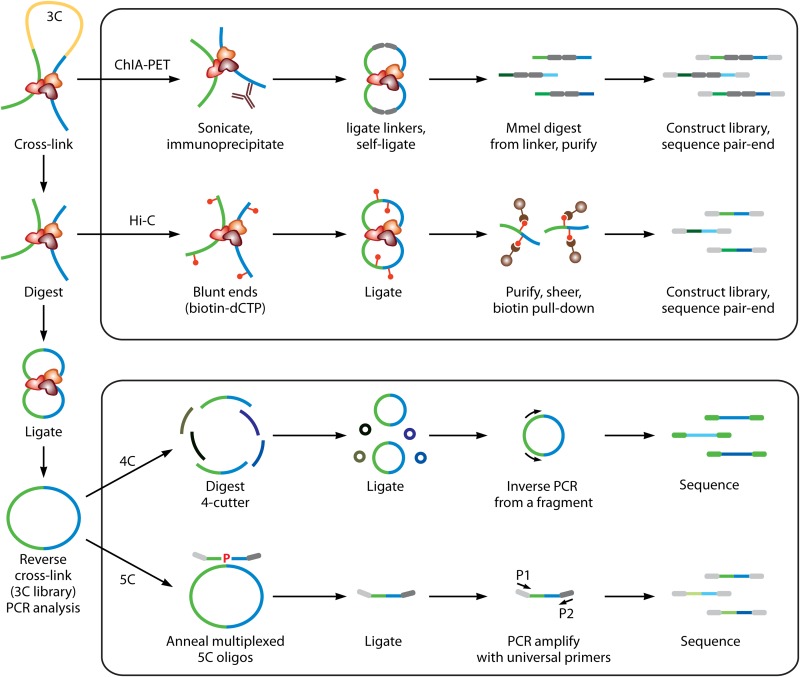
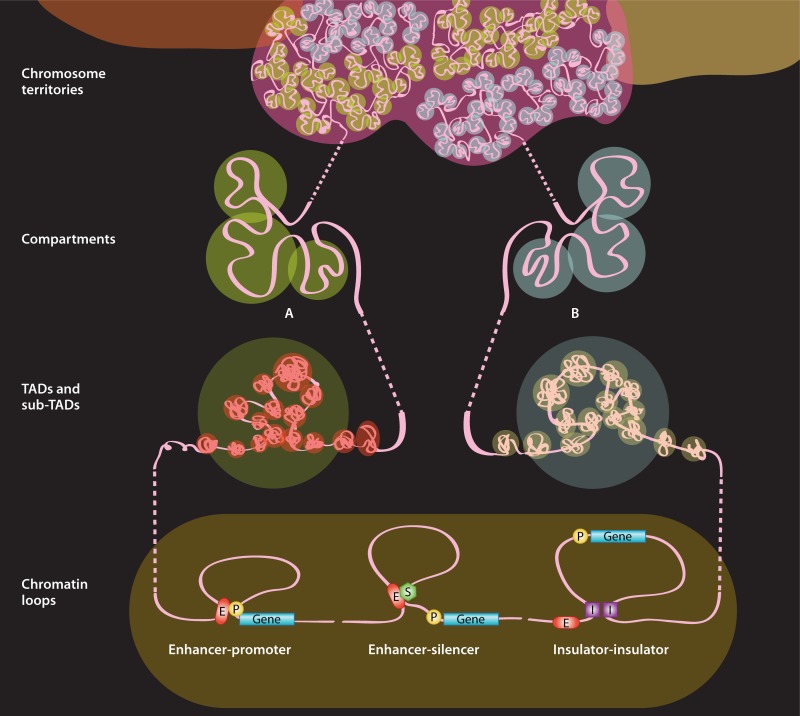
In humans, nearly two meters of genomic material must be folded to fit inside each micrometer-scale cell nucleus while remaining accessible for gene transcription, DNA replication, and DNA repair. This fact highlights the need for mechanisms governing genome organization during any activity and to maintain the physical organization of chromosomes at all times. Insight into the functions and three-dimensional structures of genomes comes mostly from the application of visual techniques such as fluorescence in situ hybridization (FISH) and molecular approaches including chromosome conformation capture (3C) technologies. Recent developments in both types of approaches now offer the possibility of exploring the folded state of an entire genome and maybe even the identification of how complex molecular machines govern its shape. In this review, we present key methodologies used to study genome organization and discuss what they reveal about chromosome conformation as it relates to transcription regulation across genomic scales in mammals.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures




Similar articles
-
Exploring the three-dimensional organization of genomes: interpreting chromatin interaction data.Nat Rev Genet. 2013 Jun;14(6):390-403. doi: 10.1038/nrg3454. Epub 2013 May 9. Nat Rev Genet. 2013. PMID: 23657480 Free PMC article. Review.
-
Normalization of a chromosomal contact map.BMC Genomics. 2012 Aug 30;13:436. doi: 10.1186/1471-2164-13-436. BMC Genomics. 2012. PMID: 22935139 Free PMC article.
-
3D genome structure modeling by Lorentzian objective function.Nucleic Acids Res. 2017 Feb 17;45(3):1049-1058. doi: 10.1093/nar/gkw1155. Nucleic Acids Res. 2017. PMID: 28180292 Free PMC article.
-
Unravelling global genome organization by 3C-seq.Semin Cell Dev Biol. 2012 Apr;23(2):213-21. doi: 10.1016/j.semcdb.2011.11.003. Epub 2011 Nov 18. Semin Cell Dev Biol. 2012. PMID: 22120510 Free PMC article. Review.
-
A maximum likelihood algorithm for reconstructing 3D structures of human chromosomes from chromosomal contact data.BMC Genomics. 2018 Feb 23;19(1):161. doi: 10.1186/s12864-018-4546-8. BMC Genomics. 2018. PMID: 29471801 Free PMC article.
Cited by
-
Nonequilibrium Biophysical Processes Influence the Large-Scale Architecture of the Cell Nucleus.Biophys J. 2020 May 5;118(9):2229-2244. doi: 10.1016/j.bpj.2019.11.017. Epub 2019 Nov 22. Biophys J. 2020. PMID: 31818465 Free PMC article.
-
The Talented LncRNAs: Meshing into Transcriptional Regulatory Networks in Cancer.Cancers (Basel). 2023 Jun 30;15(13):3433. doi: 10.3390/cancers15133433. Cancers (Basel). 2023. PMID: 37444543 Free PMC article. Review.
-
tRNA Genes Affect Chromosome Structure and Function via Local Effects.Mol Cell Biol. 2019 Apr 2;39(8):e00432-18. doi: 10.1128/MCB.00432-18. Print 2019 Apr 15. Mol Cell Biol. 2019. PMID: 30718362 Free PMC article.
-
DNA Supercoiling, Topoisomerases, and Cohesin: Partners in Regulating Chromatin Architecture?Int J Mol Sci. 2018 Mar 16;19(3):884. doi: 10.3390/ijms19030884. Int J Mol Sci. 2018. PMID: 29547555 Free PMC article. Review.
-
The 3D Genome as Moderator of Chromosomal Communication.Cell. 2016 Mar 10;164(6):1110-1121. doi: 10.1016/j.cell.2016.02.007. Cell. 2016. PMID: 26967279 Free PMC article. Review.
References
-
- Avery OT, Macleod CM, McCarty M. 1944. Studies on the chemical nature of the substance inducing transformation of pneumococcal types: induction of transformation by a desoxyribonucleic acid fraction isolated from pneumococcus type III. J Exp Med 79:137–158. doi:10.1084/jem.79.2.137. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
