

Rheumatoid Arthritis, Immunosenescence and the Hallmarks of Aging
- PMID: 26212057
- PMCID: PMC5388800
- DOI: 10.2174/1874609808666150727110744
Rheumatoid Arthritis, Immunosenescence and the Hallmarks of Aging
Abstract
Age is the most important risk factor for the development of infectious diseases, cancer and chronic inflammatory diseases including rheumatoid arthritis (RA). The very act of living causes damage to cells. A network of molecular, cellular and physiological maintenance and repair systems creates a buffering capacity against these damages. Aging leads to progressive shrinkage of the buffering capacity and increases vulnerability. In order to better understand the complex mammalian aging processes, nine hallmarks of aging and their interrelatedness were recently put forward. RA is a chronic autoimmune disease affecting the joints. Although RA may develop at a young age, the incidence of RA increases with age. It has been suggested that RA may develop as a consequence of premature aging (immunosenescence) of the immune system. Alternatively, premature aging may be the consequence of the inflammatory state in RA. In an effort to answer this chicken and egg conundrum, we here outline and discuss the nine hallmarks of aging, their contribution to the pre-aged phenotype and the effects of treatment on the reversibility of immunosenescence in RA.
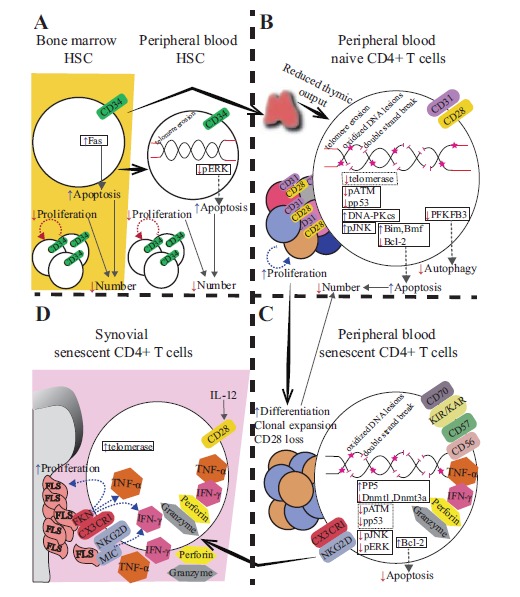
Figures
Similar articles
-
Immunosenescence of T cells: a key player in rheumatoid arthritis.Inflamm Res. 2022 Dec;71(12):1449-1462. doi: 10.1007/s00011-022-01649-0. Epub 2022 Oct 25. Inflamm Res. 2022. PMID: 36280621 Review.
-
Aging of the Immune System. Mechanisms and Therapeutic Targets.Ann Am Thorac Soc. 2016 Dec;13 Suppl 5(Suppl 5):S422-S428. doi: 10.1513/AnnalsATS.201602-095AW. Ann Am Thorac Soc. 2016. PMID: 28005419 Free PMC article. Review.
-
Rheumatoid arthritis: a role for immunosenescence?J Am Geriatr Soc. 2010 Aug;58(8):1565-75. doi: 10.1111/j.1532-5415.2010.02965.x. Epub 2010 Jul 19. J Am Geriatr Soc. 2010. PMID: 20942872 Free PMC article. Review.
-
T-cell aging in rheumatoid arthritis.Curr Opin Rheumatol. 2014 Jan;26(1):93-100. doi: 10.1097/BOR.0000000000000011. Curr Opin Rheumatol. 2014. PMID: 24296720 Free PMC article. Review.
-
Premature immunosenescence in rheumatoid arthritis and multiple sclerosis patients.Ann N Y Acad Sci. 2005 Jun;1051:255-62. doi: 10.1196/annals.1361.066. Ann N Y Acad Sci. 2005. PMID: 16126966
Cited by
-
The hallmarks of aging as a conceptual framework for health and longevity research.Front Aging. 2024 Jan 15;5:1334261. doi: 10.3389/fragi.2024.1334261. eCollection 2024. Front Aging. 2024. PMID: 38292053 Free PMC article. Review.
-
Analysis of survival rate and persistence predictors of baricitinib in real-world data from a large cohort of rheumatoid arthritis patients.Curr Res Pharmacol Drug Discov. 2024 Feb 16;6:100178. doi: 10.1016/j.crphar.2024.100178. eCollection 2024. Curr Res Pharmacol Drug Discov. 2024. PMID: 38419949 Free PMC article.
-
Difficult-to-Treat Rheumatoid Arthritis in Older Adults: Implications of Ageing for Managing Patients.Drugs Aging. 2022 Nov;39(11):841-849. doi: 10.1007/s40266-022-00976-5. Epub 2022 Sep 15. Drugs Aging. 2022. PMID: 36104655 Free PMC article.
-
Circulating DNA methylation level of CXCR5 correlates with inflammation in patients with rheumatoid arthritis.Immun Inflamm Dis. 2023 Jun;11(6):e902. doi: 10.1002/iid3.902. Immun Inflamm Dis. 2023. PMID: 37382265 Free PMC article.
-
Immunosenescence of T cells: a key player in rheumatoid arthritis.Inflamm Res. 2022 Dec;71(12):1449-1462. doi: 10.1007/s00011-022-01649-0. Epub 2022 Oct 25. Inflamm Res. 2022. PMID: 36280621 Review.
References
-
- Boots A.M., Maier A.B., Stinissen P., Masson P., Lories R.J., De Keyser F. The influence of ageing on the development and management of rheumatoid arthritis. Nat. Rev. Rheumatol. 2013;9(10):604–613. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
