

Review
doi: 10.1038/nri3865.
Epub 2015 Jul 10.
The multifaceted biology of plasmacytoid dendritic cells
Affiliations
- PMID: 26160613
- PMCID: PMC4808588
- DOI: 10.1038/nri3865
Item in Clipboard
Review
The multifaceted biology of plasmacytoid dendritic cells
Nat Rev Immunol.
2015 Aug.
Abstract
Plasmacytoid dendritic cells (pDCs) are a unique DC subset that specializes in the production of type I interferons (IFNs). pDCs promote antiviral immune responses and have been implicated in the pathogenesis of autoimmune diseases that are characterized by a type I IFN signature. However, pDCs can also induce tolerogenic immune responses. In this Review, we summarize recent progress in the field of pDC biology, focusing on the molecular mechanisms that regulate the development and functions of pDCs, the pathways involved in their sensing of pathogens and endogenous nucleic acids, their functions at mucosal sites, and their roles in infection, autoimmunity and cancer.
Figures
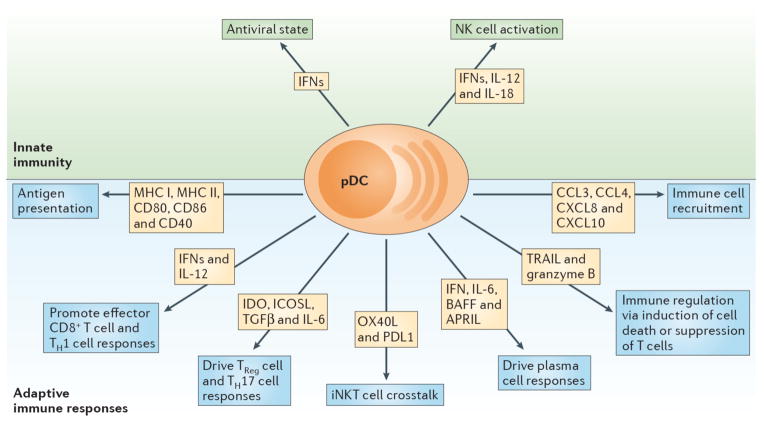
pDCs are a prominent link between innate and adaptive immune responses. Their ability to rapidly produce type I IFNs (IFN-Is) during viral infections promotes an antiviral state by inducing expression of interferon stimulated genes and apoptosis of infected cells. Moreover, IFN-I, IL-12 and IL-18 enhance NK cell activation and effector functions such as IFN-γ secretion and lysis of target cells. Expression of MHC class I (MHCI) and MHC class II (MHCII) along with costimulatory markers including CD80, CD86 and CD40 enable pDCs to cross prime CD8+ T cells and present Ag to CD4+ T cells. Production of IFN-I and IL-12 by pDCs supports accumulation and effector functions of CD8+ T cells as well as the polarization of CD4+ T cells into T helper 1 cells (Th1). pDC expression of IDO and ICOSL or production of TGF-β and IL-6 promotes TReg or Th17 commitment, respectively. Crosstalk between pDCs and invariant NKT (iNKT) cells occurs via OX40/OX40L and PD-1/PD-L1 interactions and dampens antiviral adaptive immune responses. pDCs impact B cell activation, plasma cell generation and antibody secretion through production of IFN-I, IL-6, BAFF and APRIL. TRAIL and Granzyme B (GrB) serve as immunoregulatory factors that endow pDCs with the capacity to kill tumor cells, induce apoptosis of infected CD4+ T cells and suppress T cell proliferation. Finally, pDCs secrete chemokines such CXCL8, CXCL10, CCL3 and CCL4, which attract immune cells to sites of infection or inflammation.
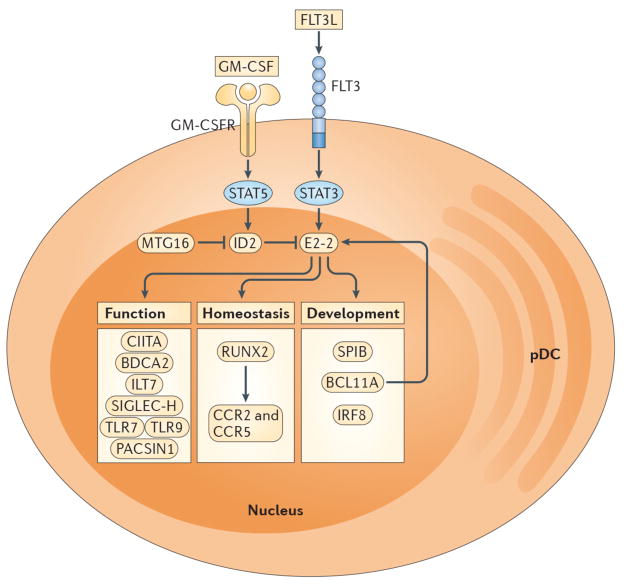
The development of pDCs from CDPs is regulated by specific cytokines and transcription factors. FLT3L-STAT3 signaling promotes expression of E2-2, which is the master transcription factor required for pDC development. The absence of E2-2 or its deletion in mature pDCs results in the complete loss of pDCs or the differentiation of pDC-committed cells into cDC-like cells, respectively. In contrast, GM-CSF-STAT5 signaling blocks pDC differentiation during DC development by inducing expression of ID2, which is an antagonist of E2-2. The transcriptional cofactor MTG16 promotes pDC differentiation and restricts cDC development in part by repressing ID2. Transcriptional targets of E2-2 encode proteins associated with pDC development, homeostasis and function. SPIB, BCL11A and IRF8 are necessary for pDC differentiation and/or survival while RUNX2 controls pDC homeostasis through expression of the chemokine receptors CCR2 and CCR5, which permit egress from the bone marrow. CIITA promotes MHC class II expression. BDCA2, ILT7 and SIGLEC-H are markers that are selectively expressed by human or mouse pDCs and are involved in regulation of type I IFN production. TLR7, TLR9 and PACSIN1 enable pDC recognition of nucleic acids and pathogens (i.e. RNA and DNA viruses), resulting in type I IFN secretion and/or pro-inflammatory cytokine production.
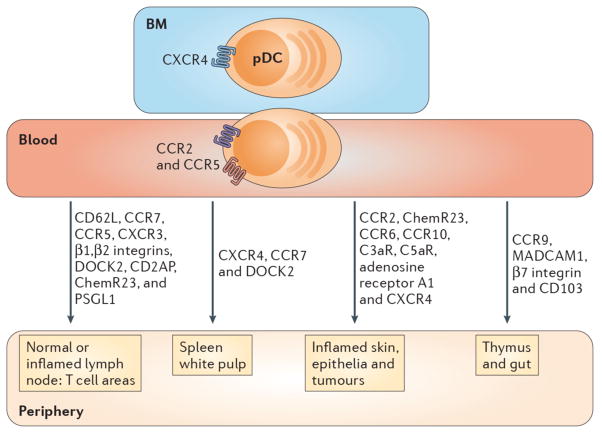
pDCs express chemokine receptors and homing molecules that promote recruitment in the steady-state and during inflammation. Development of pDCs in bone marrow (BM) stromal cell niche requires CXCR4 expression, while pDC egress from the BM into the blood is dependent on CCR5 and CCR2. pDCs are attracted to tumors that produce CXCL12 (not shown) and the splenic white pulp via CXCR4. DOCK2, a hematopoietic cell-specific CDM family protein involved in CXCR4 signaling, is also necessary for pDC migration to spleen and lymph nodes (LNs). pDCs express CD62L, PSGL-1, β1/β2 integrins and the chemokine receptors CCR5, CXCR3 and CCR7 which mediate adhesion and chemotaxis to peripheral LNs and the splenic white pulp under normal and/or inflammatory conditions. Blood pDCs express the chemerin receptor, ChemR23, as well as A1-R, C3aR and C5aR, which may guide them to peripheral LNs and damaged tissues. CD2AP is an intracellular protein that regulates actin dynamics and promotes pDC migration to LNs under inflammatory settings. CCR2 drives the recruitment of pDCs to the skin following inflammation induced by the TLR7 agonist, Imiquimod. CCR6 and CCR10 are expressed by a subset of human tonsil pDCs and enable migration to inflamed epithelia producing CCL20 and CCL27 (not shown). CCR9 and its ligand CCL25 (not shown) promote trafficking of peripheral pDCs to the thymus and are required for pDC recruitment to the small intestine under both normal and inflammatory conditions. MAdCAM-1, β7 integrin and CD103 also influence pDC trafficking to the gut. Finally, pDCs express CX3CR1 (not shown), which may impact their homeostasis.
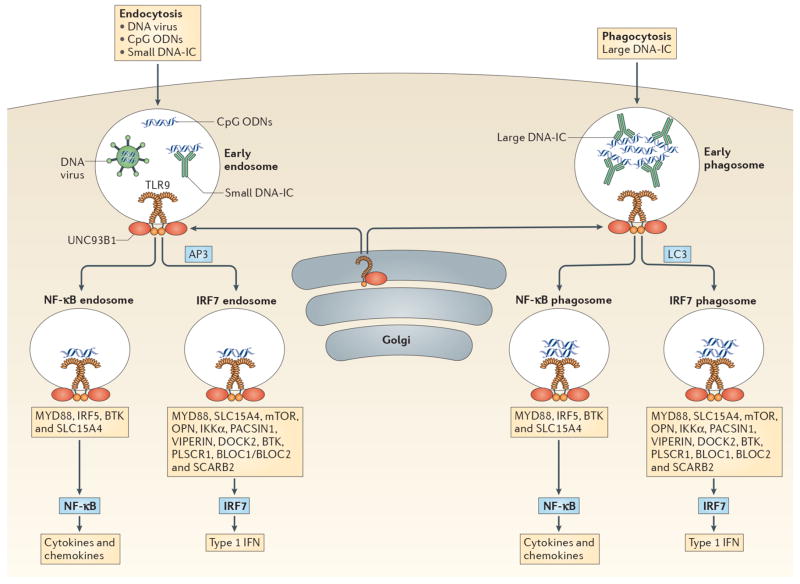
pDCs sense DNA viruses, synthetic CpG oligodeoxyribonucleotides (ODN) and endogenous DNA through TLR9. All TLR9 signaling requires MyD88 but additional factors determine whether TLR9 engagement will result in type I IFN (IFN-I) or pro-inflammatory cytokine production. These factors include mode of ligand entry, and the intracellular compartment where TLR9 encounters its ligand. TLR9 is transported to appropriate intracellular compartments by UNC93B, and requires cleavage in order to recruit MyD88,. DNA viruses, CpG ODN and small DNA immunecomplexes (DNA-ICs) enter pDCs through endocytosis and meet TLR9 in the early endosome. In contrast, large DNA-ICs are internalized by phagocytosis and encounter TLR9 and UNC93B in the early phagosome. If AP3 or LC3 are recruited then the IRF7 endosome or IRF7 phagosome is formed which leads to IFN-I production. Several molecules/pathways are involved in this process including IKKα, osteopontin (OPN), SLC15A4, BTK, BLOC1, BLOC2, DOCK2, PACSIN1, PLSCR1, VIPERIN, SCARB2 and the mTOR pathway. Alternatively, TLR9-containing compartments can form NF-κB endosomes or NF-κB phagosomes resulting in the production of pro-inflammatory cytokines and chemokines (i.e. IL-6, IL-12, TNF-α, etc). This process requires IRF5, BTK and SLC15A4. CpG ODN have different structures which results in trafficking to different compartments: CpG-A is transported to the IRF7 endosome and is a strong inducer of IFN-I, while CpG-B aggregates in the NF-κB endosome and is a potent stimulator of maturation and cytokine/chemokine production. CpG-C exhibits properties of both CpG-A and CpG-B in that it can induce both IFN-I and pro-inflammatory cytokines. TLR7 stimulation by viral and endogenous RNA may follow similar pathways.
Similar articles
-
Unraveling the functions of plasmacytoid dendritic cells during viral infections, autoimmunity, and tolerance.Immunol Rev. 2010 Mar;234(1):142-62. doi: 10.1111/j.0105-2896.2009.00881.x. Immunol Rev. 2010. PMID: 20193017 Free PMC article. Review.
-
Plasmacytoid dendritic cells: sensing nucleic acids in viral infection and autoimmune diseases.Nat Rev Immunol. 2008 Aug;8(8):594-606. doi: 10.1038/nri2358. Nat Rev Immunol. 2008. PMID: 18641647 Review.
-
Plasmacytoid dendritic cells in antiviral immunity and autoimmunity.Sci China Life Sci. 2010 Feb;53(2):172-82. doi: 10.1007/s11427-010-0045-0. Epub 2010 Mar 7. Sci China Life Sci. 2010. PMID: 20596824 Free PMC article. Review.
-
Type I Interferon Production of Plasmacytoid Dendritic Cells under Control.Int J Mol Sci. 2021 Apr 18;22(8):4190. doi: 10.3390/ijms22084190. Int J Mol Sci. 2021. PMID: 33919546 Free PMC article. Review.
-
Plasmacytoid Dendritic Cells: Development, Regulation, and Function.Immunity. 2019 Jan 15;50(1):37-50. doi: 10.1016/j.immuni.2018.12.027. Immunity. 2019. PMID: 30650380 Free PMC article. Review.
Cited by
-
Engineering dendritic cell biomimetic membrane as a delivery system for tumor targeted therapy.J Nanobiotechnology. 2024 Oct 27;22(1):663. doi: 10.1186/s12951-024-02913-7. J Nanobiotechnology. 2024. PMID: 39465376 Free PMC article. Review.
-
The TNFSF Members APRIL and BAFF and Their Receptors TACI, BCMA, and BAFFR in Oncology, With a Special Focus in Breast Cancer.Front Oncol. 2020 Jun 16;10:827. doi: 10.3389/fonc.2020.00827. eCollection 2020. Front Oncol. 2020. PMID: 32612943 Free PMC article.
-
Innate Immune Responses to Highly Pathogenic Coronaviruses and Other Significant Respiratory Viral Infections.Front Immunol. 2020 Aug 18;11:1979. doi: 10.3389/fimmu.2020.01979. eCollection 2020. Front Immunol. 2020. PMID: 32973803 Free PMC article. Review.
-
Effects of Heat-Killed Lacticaseibacillus paracasei MCC1849 on Immune Parameters in Healthy Adults-A Randomized, Double-Blind, Placebo-Controlled, Parallel-Group Study.Nutrients. 2024 Jan 9;16(2):216. doi: 10.3390/nu16020216. Nutrients. 2024. PMID: 38257109 Free PMC article. Clinical Trial.
-
Gut-Joint Axis: Impact of Bifidobacterial Cell Wall Lipoproteins on Arthritis Development.Nutrients. 2023 Nov 21;15(23):4861. doi: 10.3390/nu15234861. Nutrients. 2023. PMID: 38068720 Free PMC article.
References
-
- Lennert K, Remmele W. Karyometric research on lymph node cells in man. I. Germinoblasts, lymphoblasts & lymphocytes. Acta Haematol. 1958;19:99–113. - PubMed
-
- Siegal FP, et al. The nature of the principal type 1 interferon-producing cells in human blood. Science. 1999;284:1835–1837. - PubMed
-
- Cella M, et al. Plasmacytoid monocytes migrate to inflamed lymph nodes and produce large amounts of type I interferon. Nat Med. 1999;5:919–923. - PubMed
-
- Howell DM, Feldman M, Siegal FP, Pettera L, Fitzgerald-Bocarsly P. Peripheral blood of AIDS patients contains cells capable of providing accessory function for the natural killer cell-mediated, lysis of herpes simplex virus-infected targets despite low interferon-alpha production. J Acquir Immune Defic Syndr. 1993;6:15–23. - PubMed
-
- Perussia B, Fanning V, Trinchieri G. A leukocyte subset bearing HLA-DR antigens is responsible for in vitro alpha interferon production in response to viruses. Nat Immun Cell Growth Regul. 1985;4:120–137. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
