

Chromosome domain architecture and dynamic organization of the fission yeast genome
- PMID: 26096785
- PMCID: PMC4598268
- DOI: 10.1016/j.febslet.2015.06.008
Chromosome domain architecture and dynamic organization of the fission yeast genome
Abstract
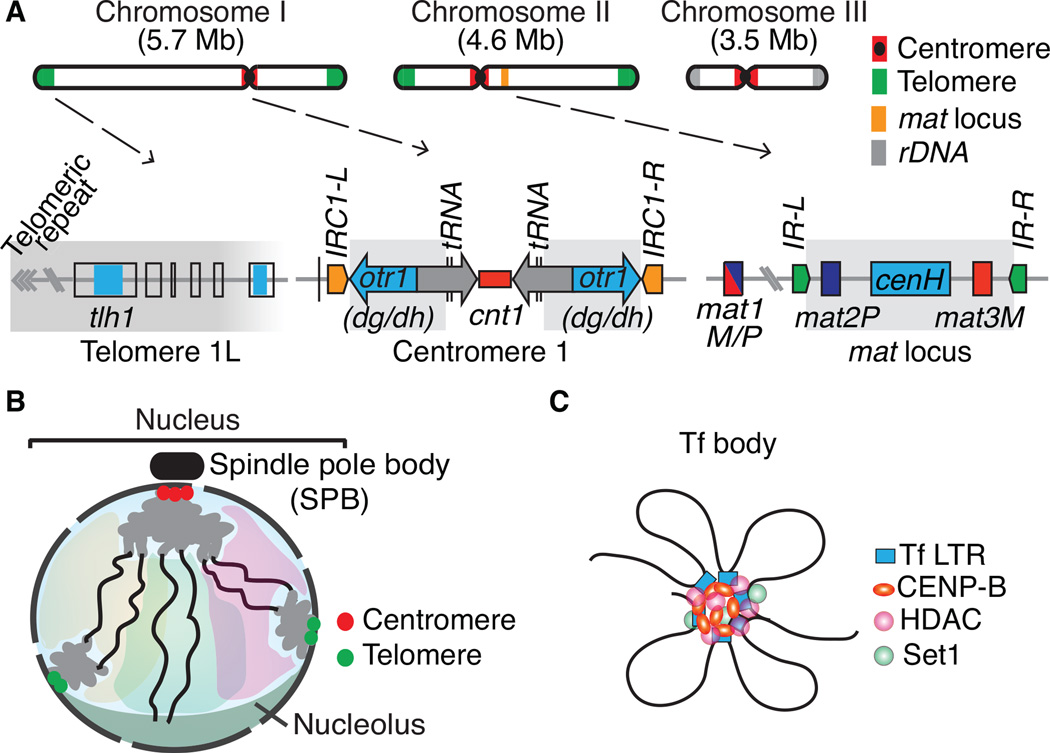
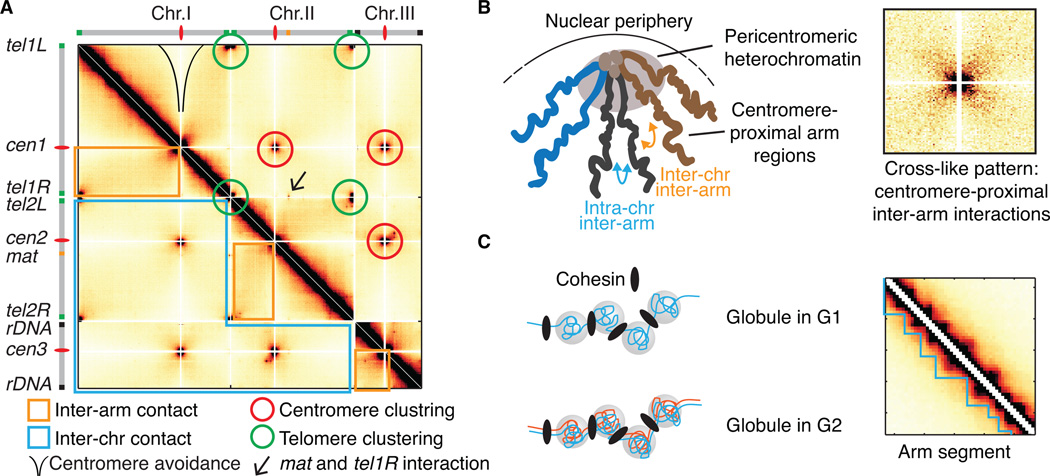
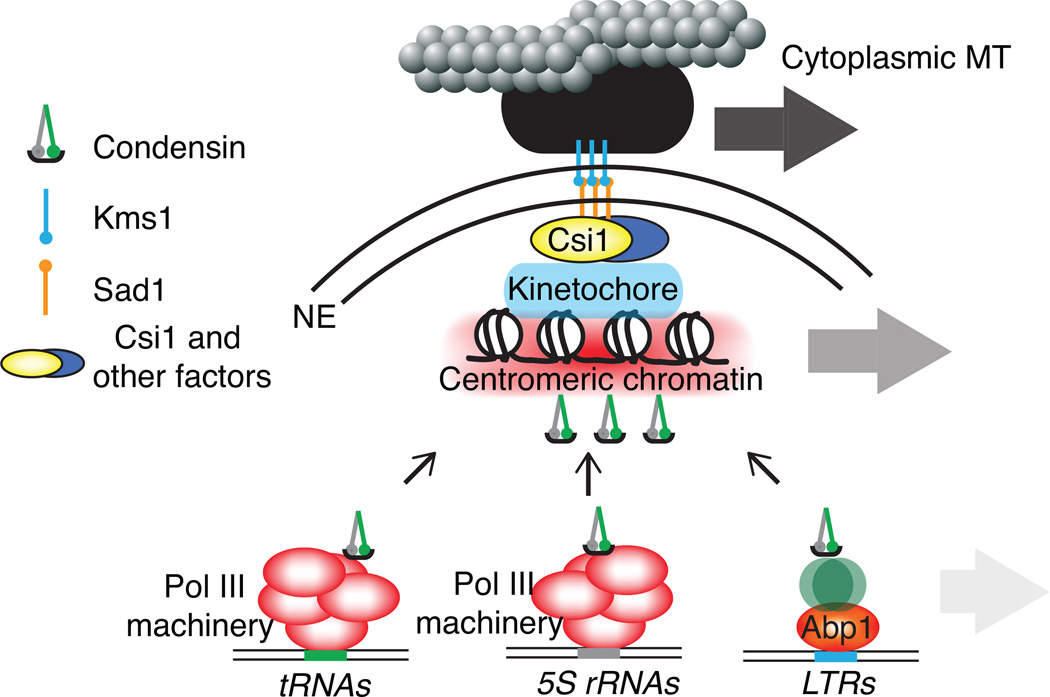
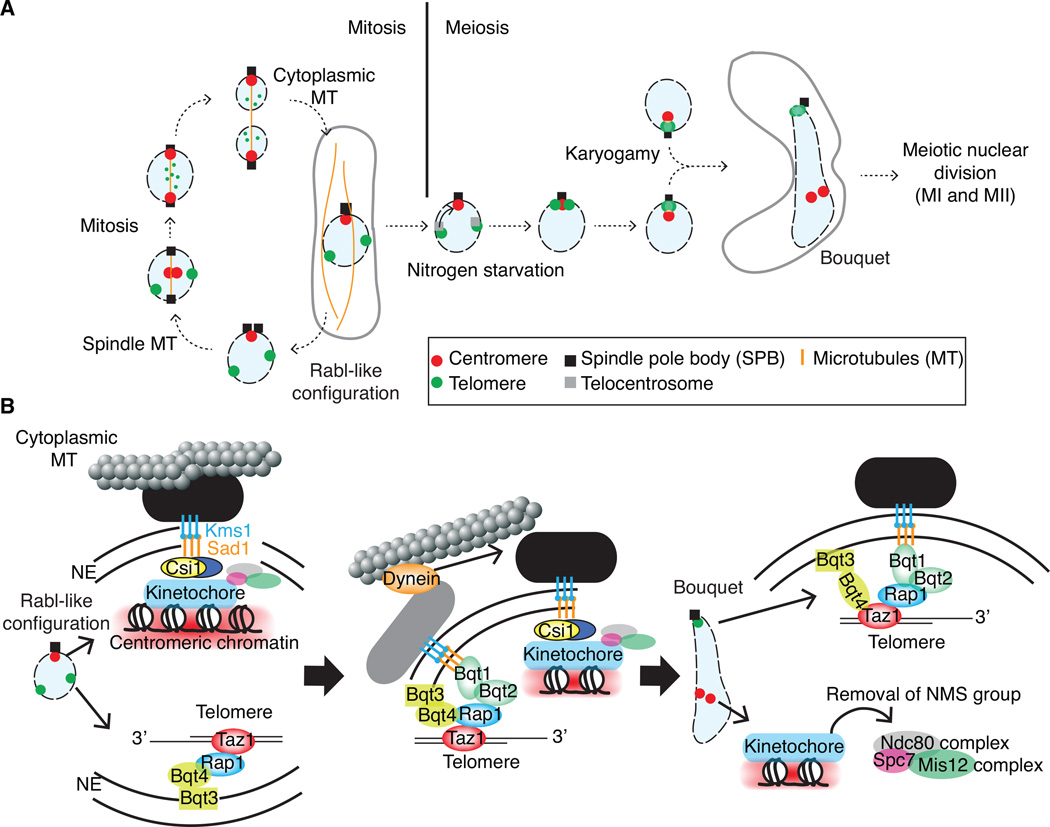
Advanced techniques including the chromosome conformation capture (3C) methodology and its derivatives are complementing microscopy approaches to study genome organization, and are revealing new details of three-dimensional (3D) genome architecture at increasing resolution. The fission yeast Schizosaccharomyces pombe (S. pombe) comprises a small genome featuring organizational elements of more complex eukaryotic systems, including conserved heterochromatin assembly machinery. Here we review key insights into genome organization revealed in this model system through a variety of techniques. We discuss the predominant role of Rabl-like configuration for interphase chromosome organization and the dynamic changes that occur during mitosis and meiosis. High resolution Hi-C studies have also revealed the presence of locally crumpled chromatin regions called "globules" along chromosome arms, and implicated a critical role for pericentromeric heterochromatin in imposing fundamental constraints on the genome to maintain chromosome territoriality and stability. These findings have shed new light on the connections between genome organization and function. It is likely that insights gained from the S. pombe system will also broadly apply to higher eukaryotes.
Keywords: Cohesin; Fission yeast; Genome organization; Heterochromatin; Hi-C; Rabl.
Published by Elsevier B.V.
Figures
Similar articles
-
Far Posterior Approach for Rib Fracture Fixation: Surgical Technique and Tips.JBJS Essent Surg Tech. 2024 Dec 6;14(4):e23.00094. doi: 10.2106/JBJS.ST.23.00094. eCollection 2024 Oct-Dec. JBJS Essent Surg Tech. 2024. PMID: 39650795 Free PMC article.
-
Depressing time: Waiting, melancholia, and the psychoanalytic practice of care.In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. PMID: 36137063 Free Books & Documents. Review.
-
Polycomb-mediated repression of paternal chromosomes maintains haploid dosage in diploid embryos of Marchantia.Elife. 2022 Aug 23;11:e79258. doi: 10.7554/eLife.79258. Elife. 2022. PMID: 35996955 Free PMC article.
-
PfMORC protein regulates chromatin accessibility and transcriptional repression in the human malaria parasite, Plasmodium falciparum.Elife. 2024 Dec 5;12:RP92499. doi: 10.7554/eLife.92499. Elife. 2024. PMID: 39636094 Free PMC article.
-
Valley fever under a changing climate in the United States.Environ Int. 2024 Nov;193:109066. doi: 10.1016/j.envint.2024.109066. Epub 2024 Oct 11. Environ Int. 2024. PMID: 39432997 Review.
Cited by
-
A stress-blinded Atf1 can fully assemble heterochromatin in a RNAi-independent minimal mat locus but impairs directionality of mat2/3 switching.iScience. 2022 Aug 2;25(8):104820. doi: 10.1016/j.isci.2022.104820. eCollection 2022 Aug 19. iScience. 2022. PMID: 35992058 Free PMC article.
-
S. pombe placed on the prion map.Microb Cell. 2017 Feb 3;4(2):35-37. doi: 10.15698/mic2017.02.555. Microb Cell. 2017. PMID: 28357387 Free PMC article.
-
The Rabl chromosome configuration masks a kinetochore reassembly mechanism in yeast mitosis.Mol Biol Cell. 2022 May 1;33(5):br8. doi: 10.1091/mbc.E20-09-0600. Epub 2022 Mar 11. Mol Biol Cell. 2022. PMID: 35274979 Free PMC article.
-
Nuclear Mechanics in the Fission Yeast.Cells. 2019 Oct 20;8(10):1285. doi: 10.3390/cells8101285. Cells. 2019. PMID: 31635174 Free PMC article. Review.
-
Computer vision for pattern detection in chromosome contact maps.Nat Commun. 2020 Nov 16;11(1):5795. doi: 10.1038/s41467-020-19562-7. Nat Commun. 2020. PMID: 33199682 Free PMC article.
References
-
- Lanctot C, Cheutin T, Cremer M, Cavalli G, Cremer T. Dynamic genome architecture in the nuclear space: regulation of gene expression in three dimensions. Nat. Rev. Genet. 2007;8:104–115. - PubMed
-
- Bickmore WA, van Steensel B. Genome architecture: domain organization of interphase chromosomes. Cell. 2013;152:1270–1284. - PubMed
-
- Nagai S, Heun P, Gasser SM. Roles for nuclear organization in the maintenance of genome stability. Epigenomics. 2010;2:289–305. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
