

Arabidopsis microRNA expression regulation in a wide range of abiotic stress responses
- PMID: 26089831
- PMCID: PMC4454879
- DOI: 10.3389/fpls.2015.00410
Arabidopsis microRNA expression regulation in a wide range of abiotic stress responses
Abstract
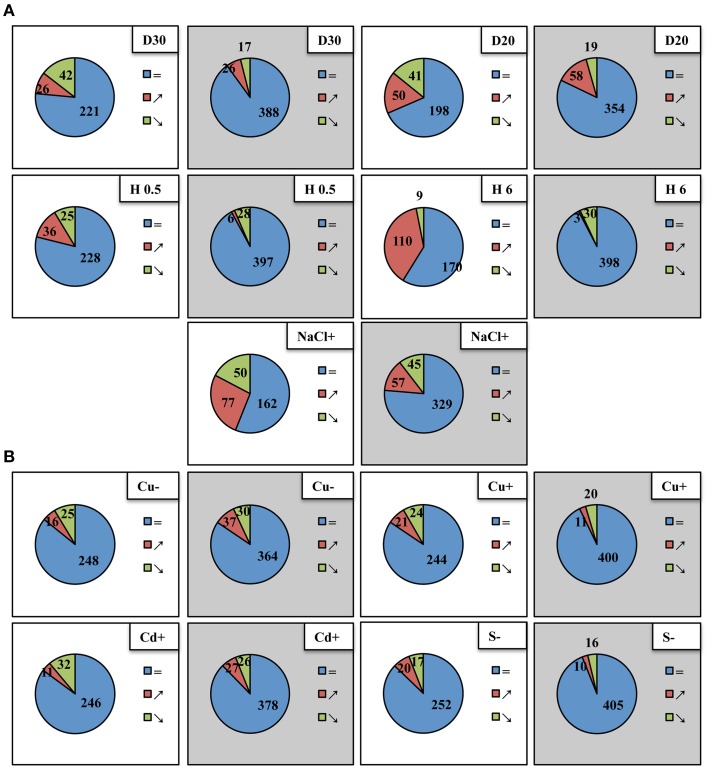
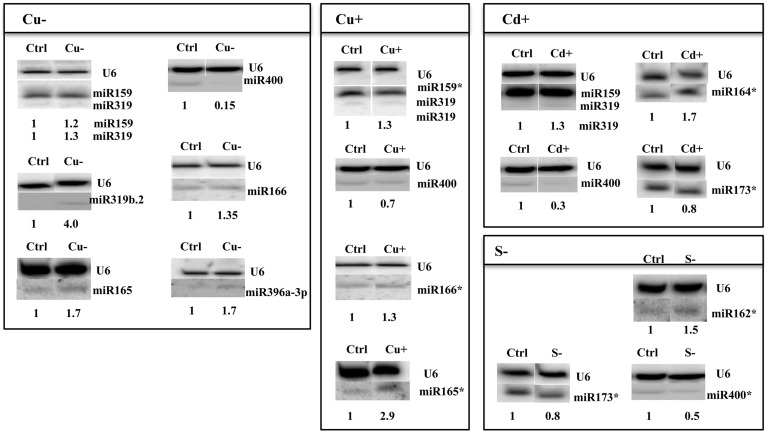
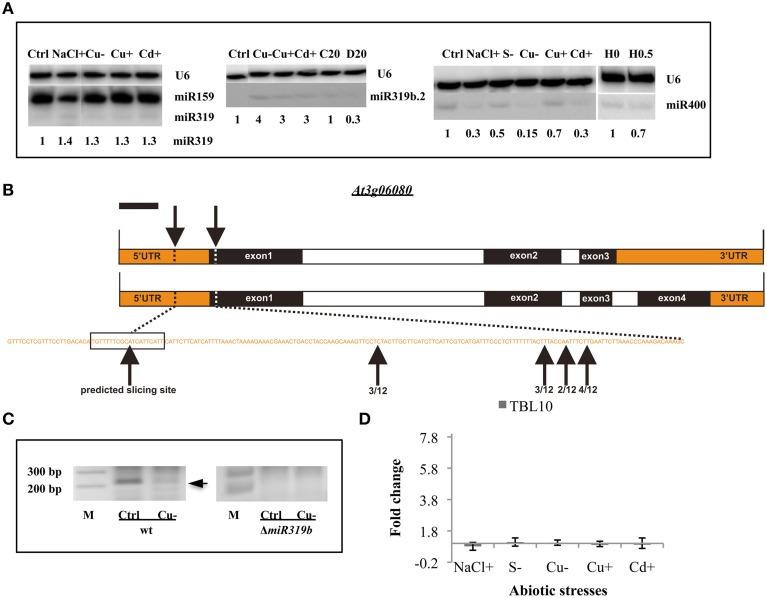
Arabidopsis microRNA expression regulation was studied in a wide array of abiotic stresses such as drought, heat, salinity, copper excess/deficiency, cadmium excess, and sulfur deficiency. A home-built RT-qPCR mirEX platform for the amplification of 289 Arabidopsis microRNA transcripts was used to study their response to abiotic stresses. Small RNA sequencing, Northern hybridization, and TaqMan® microRNA assays were performed to study the abundance of mature microRNAs. A broad response on the level of primary miRNAs (pri-miRNAs) was observed. However, stress response at the level of mature microRNAs was rather confined. The data presented show that in most instances, the level of a particular mature miRNA could not be predicted based on the level of its pri-miRNA. This points to an essential role of posttranscriptional regulation of microRNA expression. New Arabidopsis microRNAs responsive to abiotic stresses were discovered. Four microRNAs: miR319a/b, miR319b.2, and miR400 have been found to be responsive to several abiotic stresses and thus can be regarded as general stress-responsive microRNA species.
Keywords: abiotic stress; gene expression; miRNA; pri-miRNA.
Figures

Similar articles
-
Quantitative Analysis of Plant miRNA Primary Transcripts.Methods Mol Biol. 2021;2170:53-77. doi: 10.1007/978-1-0716-0743-5_5. Methods Mol Biol. 2021. PMID: 32797451
-
Profiling the Abiotic Stress Responsive microRNA Landscape of Arabidopsis thaliana.Plants (Basel). 2019 Mar 10;8(3):58. doi: 10.3390/plants8030058. Plants (Basel). 2019. PMID: 30857364 Free PMC article.
-
Arabidopsis thaliana microRNA162 level is posttranscriptionally regulated via splicing and polyadenylation site selection.Acta Biochim Pol. 2016;63(4):811-816. doi: 10.18388/abp.2016_1349. Epub 2016 Oct 5. Acta Biochim Pol. 2016. PMID: 27704058
-
MicroRNA: a new target for improving plant tolerance to abiotic stress.J Exp Bot. 2015 Apr;66(7):1749-61. doi: 10.1093/jxb/erv013. Epub 2015 Feb 19. J Exp Bot. 2015. PMID: 25697792 Free PMC article. Review.
-
[The role of microRNA in abiotic stress response in plants].Mol Biol (Mosk). 2016 May-Jun;50(3):387-94. doi: 10.7868/S0026898416020105. Mol Biol (Mosk). 2016. PMID: 27414777 Review. Russian.
Cited by
-
Salt Stress Reveals a New Role for ARGONAUTE1 in miRNA Biogenesis at the Transcriptional and Posttranscriptional Levels.Plant Physiol. 2016 Sep;172(1):297-312. doi: 10.1104/pp.16.00830. Epub 2016 Jul 6. Plant Physiol. 2016. PMID: 27385819 Free PMC article.
-
Cold-responsive miRNAs and their target genes in the wild eggplant species Solanum aculeatissimum.BMC Genomics. 2017 Dec 29;18(1):1000. doi: 10.1186/s12864-017-4341-y. BMC Genomics. 2017. PMID: 29287583 Free PMC article.
-
Gene Networks Involved in Plant Heat Stress Response and Tolerance.Int J Mol Sci. 2022 Oct 9;23(19):11970. doi: 10.3390/ijms231911970. Int J Mol Sci. 2022. PMID: 36233272 Free PMC article. Review.
-
Pi-starvation induced transcriptional changes in barley revealed by a comprehensive RNA-Seq and degradome analyses.BMC Genomics. 2021 Mar 9;22(1):165. doi: 10.1186/s12864-021-07481-w. BMC Genomics. 2021. PMID: 33750301 Free PMC article.
-
PRP4KA phosphorylates SERRATE for degradation via 20S proteasome to fine-tune miRNA production in Arabidopsis.Sci Adv. 2022 Mar 25;8(12):eabm8435. doi: 10.1126/sciadv.abm8435. Epub 2022 Mar 25. Sci Adv. 2022. PMID: 35333566 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases