

Uncovering DELLA-Independent Gibberellin Responses by Characterizing New Tomato procera Mutants
- PMID: 26036254
- PMCID: PMC4498196
- DOI: 10.1105/tpc.114.132795
Uncovering DELLA-Independent Gibberellin Responses by Characterizing New Tomato procera Mutants
Abstract
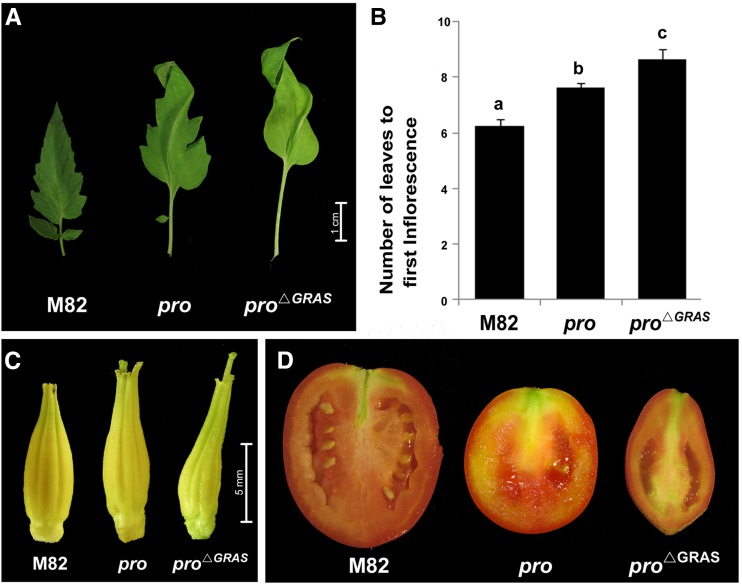
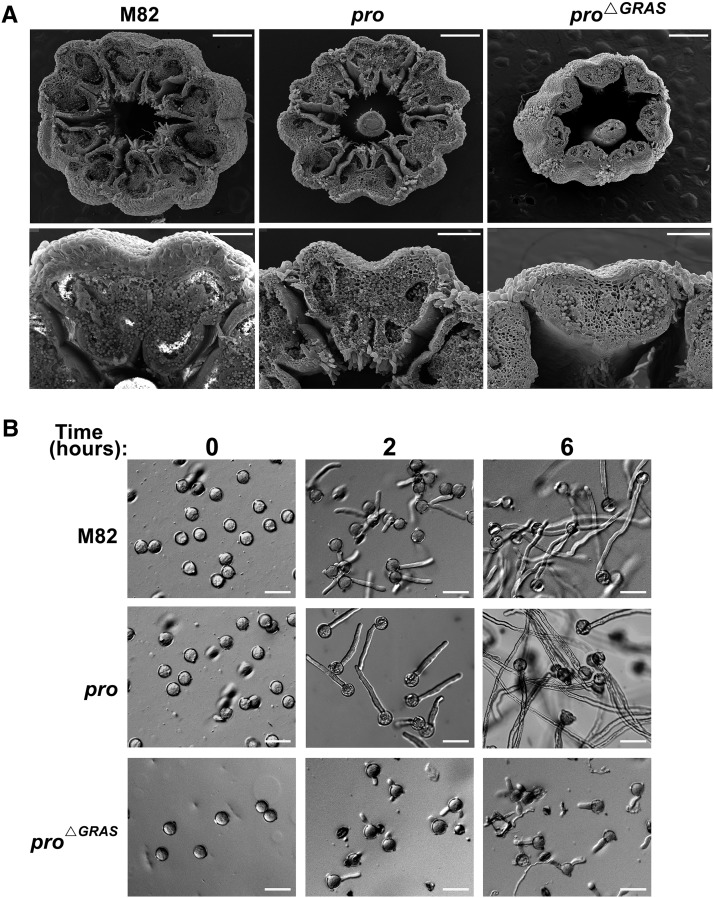
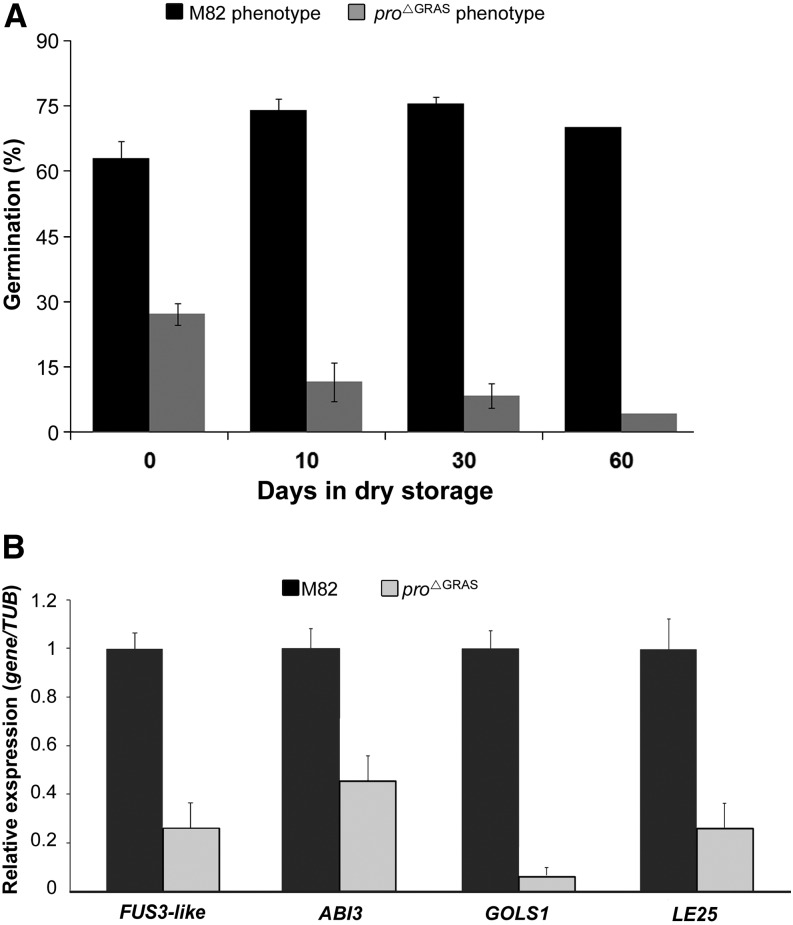
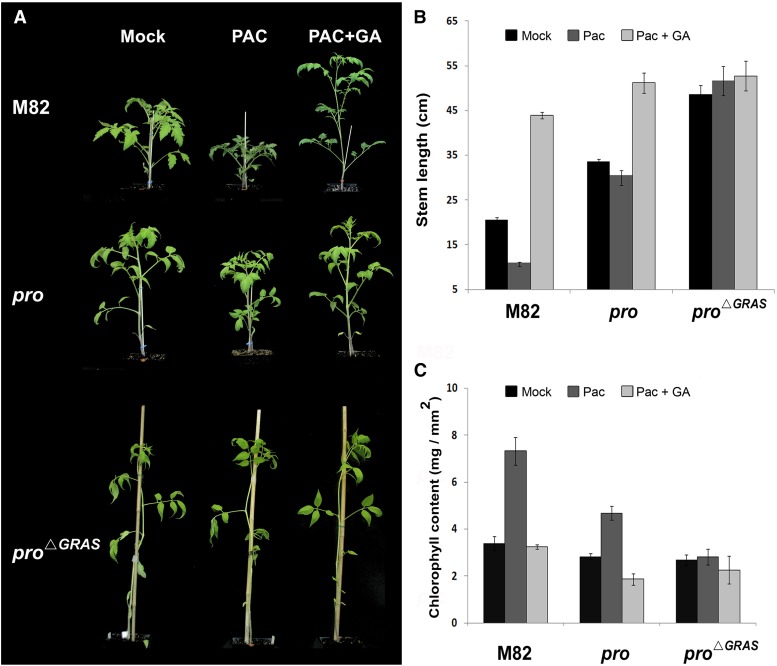
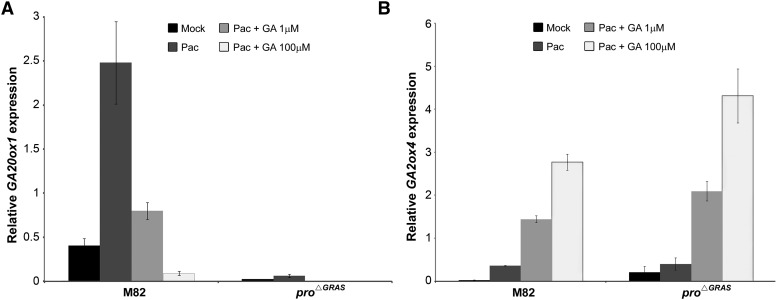
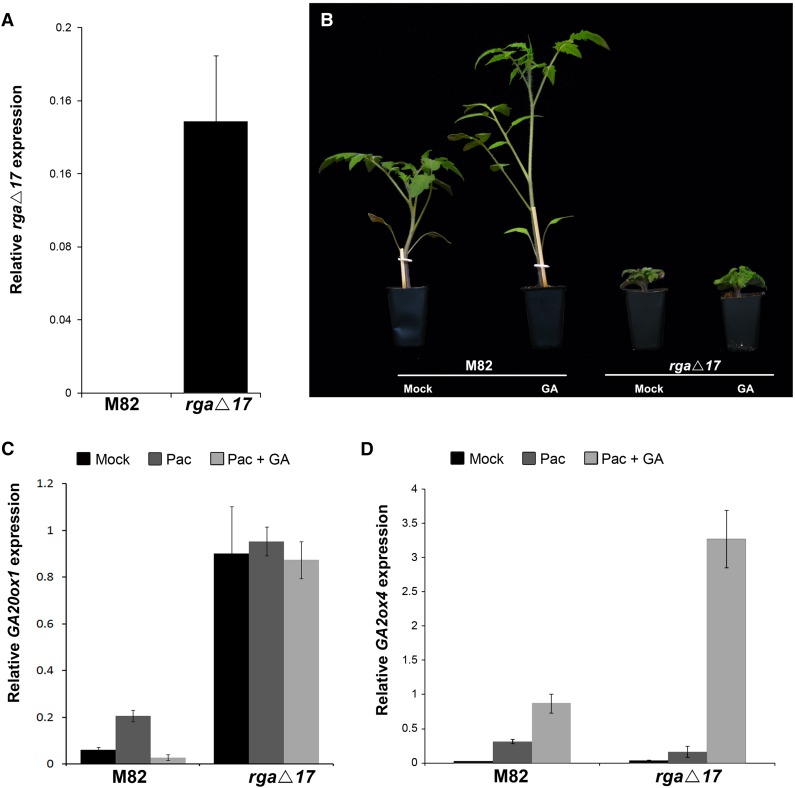
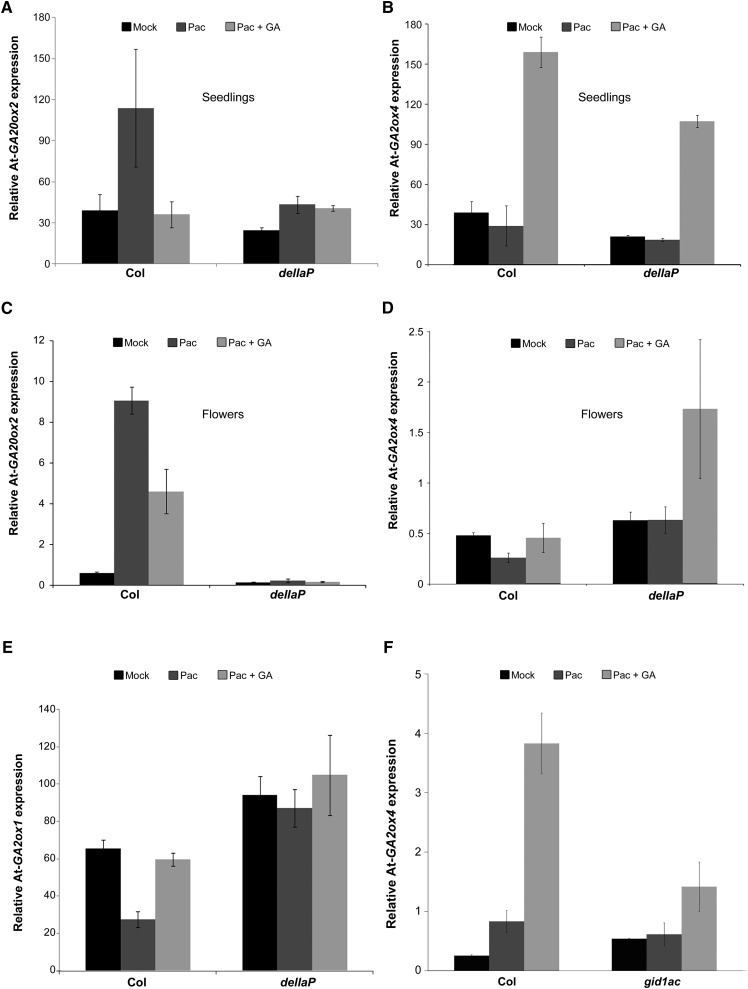
Gibberellin (GA) regulates plant development primarily by triggering the degradation/deactivation of the DELLA proteins. However, it remains unclear whether all GA responses are regulated by DELLAs. Tomato (Solanum lycopersicum) has a single DELLA gene named PROCERA (PRO), and its recessive pro allele exhibits constitutive GA activity but retains responsiveness to external GA. In the loss-of-function mutant pro(ΔGRAS), all examined GA developmental responses were considerably enhanced relative to pro and a defect in seed desiccation tolerance was uncovered. As pro, but not pro(ΔGRAS), elongation was promoted by GA treatment, pro may retain residual DELLA activity. In agreement with homeostatic feedback regulation of the GA biosynthetic pathway, we found that GA20oxidase1 expression was suppressed in pro(ΔGRAS) and was not affected by exogenous GA3. In contrast, expression of GA2oxidase4 was not affected by the elevated GA signaling in pro(ΔGRAS) but was strongly induced by exogenous GA3. Since a similar response was found in Arabidopsis thaliana plants with impaired activity of all five DELLA genes, we suggest that homeostatic GA responses are regulated by both DELLA-dependent and -independent pathways. Transcriptome analysis of GA-treated pro(ΔGRAS) leaves suggests that 5% of all GA-regulated genes in tomato are DELLA independent.
© 2015 American Society of Plant Biologists. All rights reserved.
Figures

Similar articles
-
PROCERA encodes a DELLA protein that mediates control of dissected leaf form in tomato.Plant J. 2008 Nov;56(4):603-12. doi: 10.1111/j.1365-313X.2008.03628.x. Epub 2008 Jul 16. Plant J. 2008. PMID: 18643984
-
Identification and functional study of a mild allele of SlDELLA gene conferring the potential for improved yield in tomato.Sci Rep. 2018 Aug 13;8(1):12043. doi: 10.1038/s41598-018-30502-w. Sci Rep. 2018. PMID: 30104574 Free PMC article.
-
Procera is a putative DELLA mutant in tomato (Solanum lycopersicum): effects on the seed and vegetative plant.J Exp Bot. 2008;59(3):585-93. doi: 10.1093/jxb/erm354. Epub 2008 Feb 3. J Exp Bot. 2008. PMID: 18250077
-
Gibberellin metabolism and signaling.Vitam Horm. 2005;72:289-338. doi: 10.1016/S0083-6729(05)72009-4. Vitam Horm. 2005. PMID: 16492475 Review.
-
Gibberellins and abscisic acid signal crosstalk: living and developing under unfavorable conditions.Plant Cell Rep. 2013 Jul;32(7):1007-16. doi: 10.1007/s00299-013-1409-2. Epub 2013 Mar 23. Plant Cell Rep. 2013. PMID: 23525744 Review.
Cited by
-
Fruit setting rewires central metabolism via gibberellin cascades.Proc Natl Acad Sci U S A. 2020 Sep 22;117(38):23970-23981. doi: 10.1073/pnas.2011859117. Epub 2020 Sep 3. Proc Natl Acad Sci U S A. 2020. PMID: 32883877 Free PMC article.
-
Potential function of CbuSPL and gene encoding its interacting protein during flowering in Catalpa bungei.BMC Plant Biol. 2020 Mar 6;20(1):105. doi: 10.1186/s12870-020-2303-z. BMC Plant Biol. 2020. PMID: 32143577 Free PMC article.
-
Using CRISPR/Cas9 genome editing in tomato to create a gibberellin-responsive dominant dwarf DELLA allele.Plant Biotechnol J. 2019 Jan;17(1):132-140. doi: 10.1111/pbi.12952. Epub 2018 Jun 22. Plant Biotechnol J. 2019. PMID: 29797460 Free PMC article.
-
Systematic Analysis of Gibberellin Pathway Components in Medicago truncatula Reveals the Potential Application of Gibberellin in Biomass Improvement.Int J Mol Sci. 2020 Sep 29;21(19):7180. doi: 10.3390/ijms21197180. Int J Mol Sci. 2020. PMID: 33003317 Free PMC article.
-
Regulatory Networks in Pollen Development under Cold Stress.Front Plant Sci. 2016 Mar 31;7:402. doi: 10.3389/fpls.2016.00402. eCollection 2016. Front Plant Sci. 2016. PMID: 27066044 Free PMC article. Review.
References
-
- Achard P., Cheng H., De Grauwe L., Decat J., Schoutteten H., Moritz T., Van Der Straeten D., Peng J., Harberd N.P. (2006). Integration of plant responses to environmentally activated phytohormonal signals. Science 311: 91–94. - PubMed
-
- Achard P., Renou J.P., Berthomé R., Harberd N.P., Genschik P. (2008). Plant DELLAs restrain growth and promote survival of adversity by reducing the levels of reactive oxygen species. Curr. Biol. 18: 656–660. - PubMed
-
- Angelovici R., Galili G., Fernie A.R., Fait A. (2010). Seed desiccation: a bridge between maturation and germination. Trends Plant Sci. 15: 211–218. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
