

Junctional actin assembly is mediated by Formin-like 2 downstream of Rac1
- PMID: 25963818
- PMCID: PMC4427798
- DOI: 10.1083/jcb.201412015
Junctional actin assembly is mediated by Formin-like 2 downstream of Rac1
Abstract
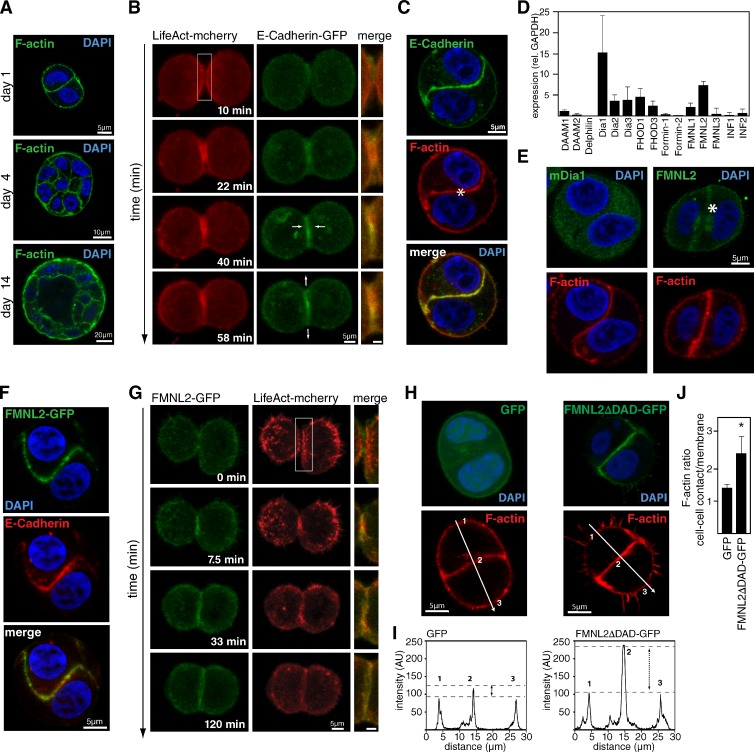
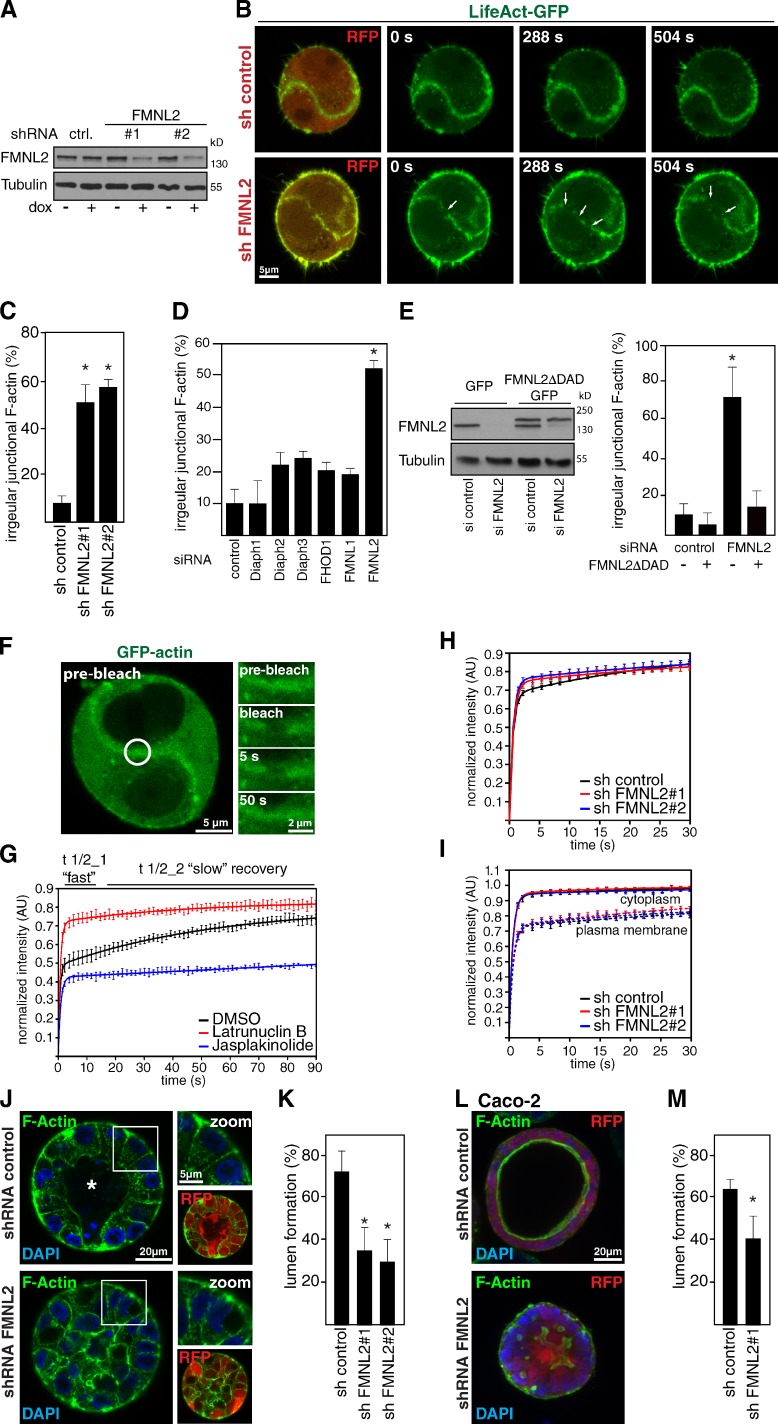
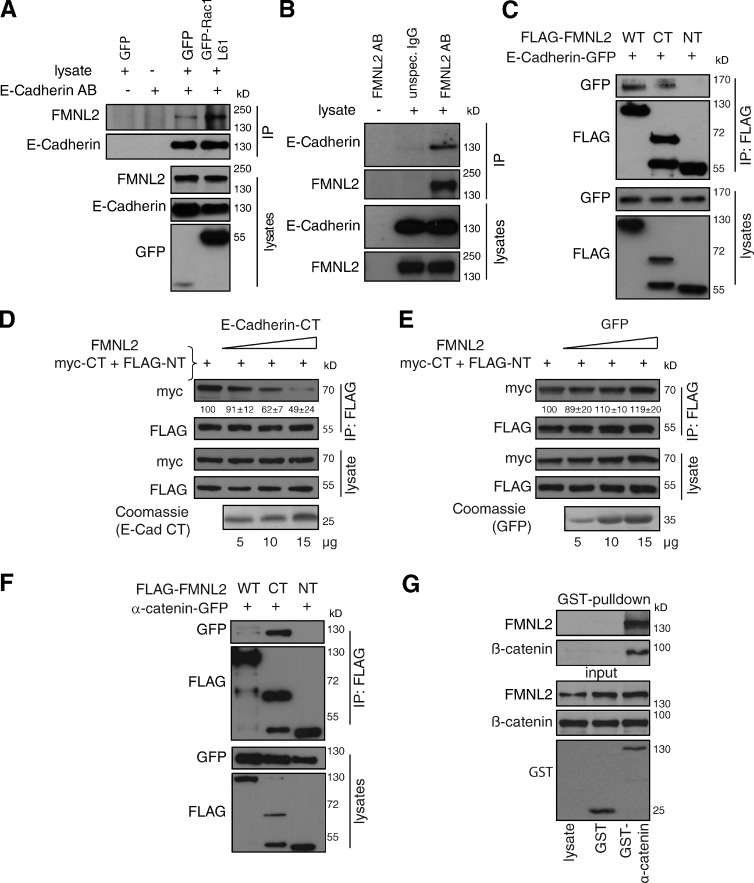
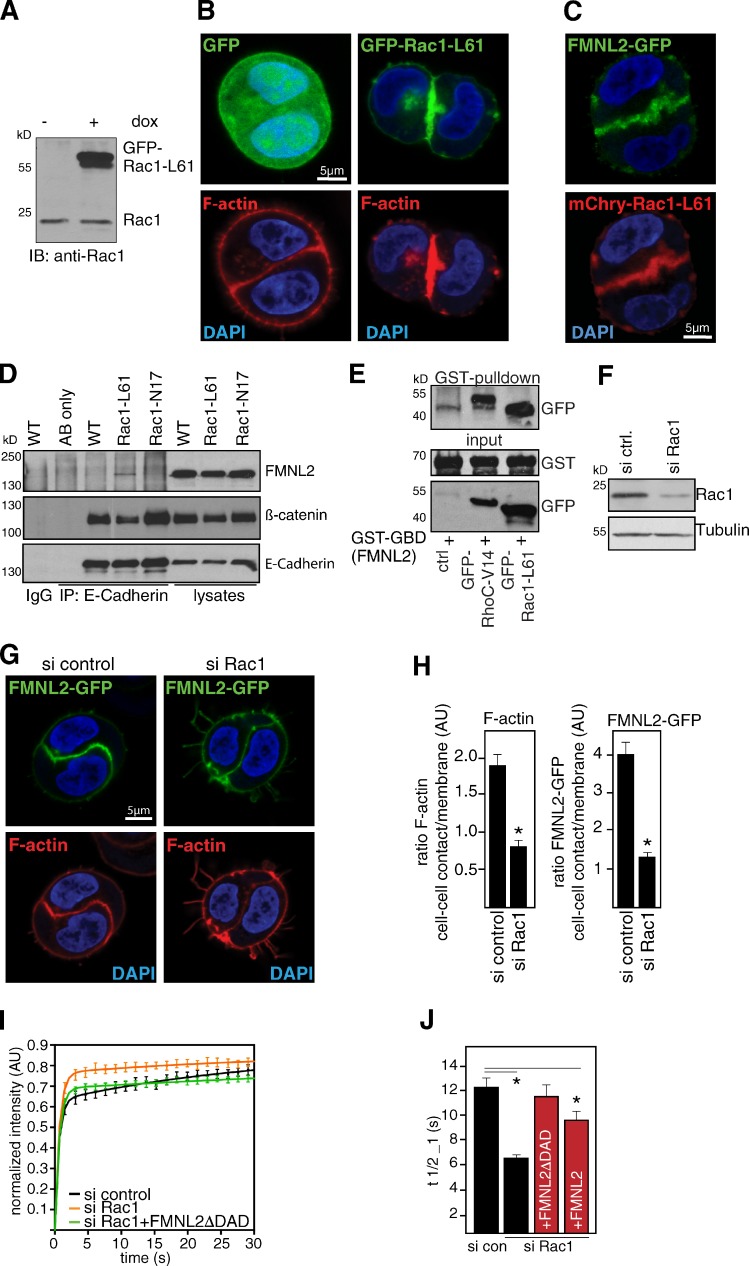
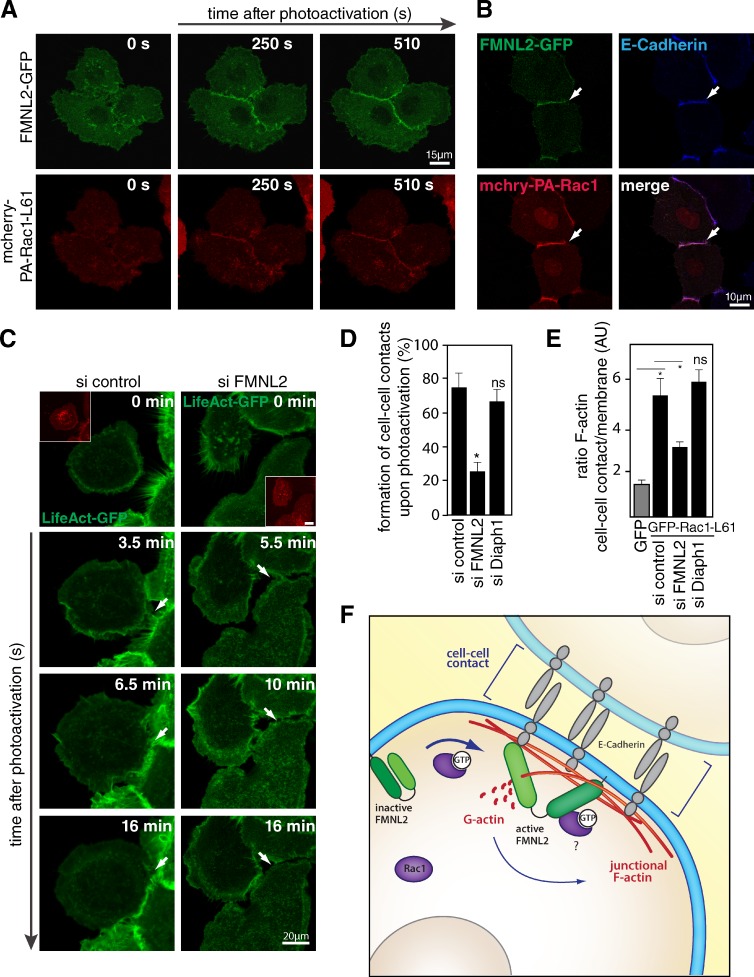
Epithelial integrity is vitally important, and its deregulation causes early stage cancer. De novo formation of an adherens junction (AJ) between single epithelial cells requires coordinated, spatial actin dynamics, but the mechanisms steering nascent actin polymerization for cell-cell adhesion initiation are not well understood. Here we investigated real-time actin assembly during daughter cell-cell adhesion formation in human breast epithelial cells in 3D environments. We identify formin-like 2 (FMNL2) as being specifically required for actin assembly and turnover at newly formed cell-cell contacts as well as for human epithelial lumen formation. FMNL2 associates with components of the AJ complex involving Rac1 activity and the FMNL2 C terminus. Optogenetic control of Rac1 in living cells rapidly drove FMNL2 to epithelial cell-cell contact zones. Furthermore, Rac1-induced actin assembly and subsequent AJ formation critically depends on FMNL2. These data uncover FMNL2 as a driver for human epithelial AJ formation downstream of Rac1.
© 2015 Grikscheit et al.
Figures
Similar articles
-
A Rac1-FMNL2 signaling module affects cell-cell contact formation independent of Cdc42 and membrane protrusions.PLoS One. 2018 Mar 26;13(3):e0194716. doi: 10.1371/journal.pone.0194716. eCollection 2018. PLoS One. 2018. PMID: 29579104 Free PMC article.
-
Differential roles for actin polymerization and a myosin II motor in assembly of the epithelial apical junctional complex.Mol Biol Cell. 2005 Jun;16(6):2636-50. doi: 10.1091/mbc.e05-01-0043. Epub 2005 Mar 30. Mol Biol Cell. 2005. PMID: 15800060 Free PMC article.
-
MEK/ERK regulates adherens junctions and migration through Rac1.Cell Motil Cytoskeleton. 2007 Mar;64(3):143-56. doi: 10.1002/cm.20172. Cell Motil Cytoskeleton. 2007. PMID: 17183546
-
Actin dynamics and cell-cell adhesion in epithelia.Curr Opin Cell Biol. 2001 Feb;13(1):76-84. doi: 10.1016/s0955-0674(00)00177-0. Curr Opin Cell Biol. 2001. PMID: 11163137 Review.
-
Formins at the Junction.Trends Biochem Sci. 2016 Feb;41(2):148-159. doi: 10.1016/j.tibs.2015.12.002. Epub 2015 Dec 28. Trends Biochem Sci. 2016. PMID: 26732401 Review.
Cited by
-
Postmitotic expansion of cell nuclei requires nuclear actin filament bundling by α-actinin 4.EMBO Rep. 2020 Nov 5;21(11):e50758. doi: 10.15252/embr.202050758. Epub 2020 Sep 22. EMBO Rep. 2020. PMID: 32959960 Free PMC article.
-
Characterization of a L136P mutation in Formin-like 2 (FMNL2) from a patient with chronic inflammatory bowel disease.PLoS One. 2021 May 27;16(5):e0252428. doi: 10.1371/journal.pone.0252428. eCollection 2021. PLoS One. 2021. PMID: 34043722 Free PMC article.
-
Phenotypic Plasticity of Cancer Cells Based on Remodeling of the Actin Cytoskeleton and Adhesive Structures.Int J Mol Sci. 2021 Feb 12;22(4):1821. doi: 10.3390/ijms22041821. Int J Mol Sci. 2021. PMID: 33673054 Free PMC article. Review.
-
A specific FMNL2 isoform is up-regulated in invasive cells.BMC Cell Biol. 2016 Aug 30;17(1):32. doi: 10.1186/s12860-016-0110-z. BMC Cell Biol. 2016. PMID: 27578625 Free PMC article.
-
The multifarious regulation of the apical junctional complex.Open Biol. 2020 Feb;10(2):190278. doi: 10.1098/rsob.190278. Epub 2020 Feb 19. Open Biol. 2020. PMID: 32070233 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
